馬科(化石)
正文
現生馬(包括斑馬、驢和半驢)從形態上講是高度特化的一類動物:牙齒高冠,肢為單趾。現生代表只有一屬,但化石馬的屬種很多,各屬之間的聯繫較易辨認。因此,達爾文進化理論問世(1859)後,馬的進化成了古生物學支持進化論的最早的例證。1861年英人R.歐文首先提出了一個進化序列:古獸馬(Palaeotherium)→安琪馬(Anchitherium)→三趾馬(Hipparion)→馬(Equus)。B.O.科瓦列夫斯基對其中的某些環節作了極為細緻的剖析,而T.M.赫胥黎對這一思想進行廣泛宣傳。達爾文主義之被普遍接受,馬的進化,作為古生物學的具說服力的實證,是起了很大作用的。但是隨著在北美愈來愈多的新的馬類化石的發現,人們逐漸認識到,馬類的進化遠非想像的那樣簡單。到1940年R.A.斯特頓發表“北美馬科系統發育”一文時,馬的進化歷史才逐漸被大體認識,並被人們接受。這一認識的主要論點,至今仍被廣泛接受和被引用。根據這一認識,馬類的進化主幹一直在北美。最早可追溯至距今約5500萬年,主幹之外,尚有若干旁支,並在不同的地質時期內擴大到其他大陸(見圖),目前公認的屬有20多個。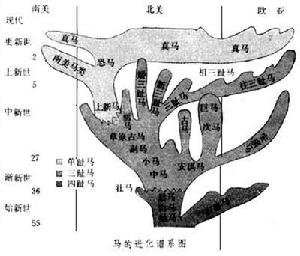 馬科(化石)
馬科(化石)