食肉目(化石)
正文
食肉目動物是指真獸或有胎盤亞綱中以捕食熱血動物為主的,包括生活於水中和陸上的兩大類哺乳動物。到上一世紀後葉人們對食肉動物已相當了解。其形態特徵主要是有由上第四前臼齒和第一下臼齒組成了一對具切割作用的裂齒,犬齒粗大,頭骨上矢狀嵴高聳,顳窩及下頜冠狀突大,以容納強壯的顳肌,具骨質耳泡,下頜關節突位置低,四肢靈活,尺、撓骨分離,腕骨中舟狀骨、月骨和頭狀骨常癒合,4或5趾,末端具銳爪。科以上的分類主要依據頭骨基部,特別是耳區附近的構造。在19世紀,人們習慣地把陸上食肉動物分成以犬和貓為代表的兩個超科。前者除犬科外還包括熊、浣熊和鼬科,後者除貓科外則包括靈貓科和鬣狗科。前者耳泡腹壁僅由內鼓骨組成,大多有翼蝶管;後者耳泡膨大分為二室,腹壁由內、外兩塊鼓骨組成,無翼蝶管。20世紀化石新材料的發現和深入研究對上述分類提出了修正和補充。首先增加了一個超科,細齒獸超科(Miacoidea)。這類動物發現於全北界的古新統和始新統中。它們雖已具有真正的裂齒,但在其他方面都還很原始,身體都很小,耳泡尚未骨化,前臼齒不退化,臼齒仍為磨楔型,腕骨中也無癒合現象。其次,發現犬科在系統關係上與熊、浣熊和鼬科都相距較遠,而與貓科反而更接近。這從耳區解剖和血清研究上都得到了證實。所以陸上食肉動物可以劃分為熊和犬貓兩個大的分類單元。水生食肉動物形態極特化,身體呈流線型,四肢漿狀,牙齒同型化並趨於消失。最初人們多把它們單列一目,但更多的事實表明它們是從陸生食肉動物中返回水中的一支。有人根據形態把它們和陸生食肉動物分列為兩個亞目:水生的鰭腳亞目(Pinnepedia)和陸上的裂腳亞目(Fissipedia)。但也有人根據分支系統學的觀點把耳海豹 (Otariidae)和海象(Odobenidae)與浣熊及熊歸在一起,而把無耳海豹(Phocidae)和鼬類歸在一起。食肉目在科一級的分類上爭議較少。分歧較大的是關於大熊貓的分類位置。有人認為它和熊類更接近。有人把它和原歸入浣熊科的小熊貓合在一起,建立熊貓科(Ailuridae)。也有人把大熊貓單獨建立一科(Ailuropo-didae)。最近從中國祿豐古猿地點發現的原始大熊貓化石表明,它還是和熊科的關係更接近。熊貓科現生種局限於東洋界的北部,最早的化石發現於全北界的上上新統。中國在上上新統至中更新統中發現的化石相當多。
浣熊科 (Procyonidae)現生種限於新大陸。這是一類小至中型的動物。它們的上裂齒都有較大的原尖和次尖。臼齒多呈方或長方形,具半封閉的外耳道上窩。化石主要發現於北美。最早出現於早漸新世。最近有人根據耳區構造將歐洲漸新統中發現的Pleisictic等也歸入這一科中。中國至今未發現過這一科的化石。
熊科 (Ursidae)現生種分布於全北界和南美洲。這是一些體型笨重的動物。它們的裂齒變小,前臼齒退縮,臼齒加大,冠面紋飾複雜化,基枕部短寬,耳泡扁平,外耳道長。它們大概起源於歐洲漸新世的鼬熊(Cephalogale)。晚第三紀的主要代表是祖熊(Ursavus),這個屬在北美和中國也已發現,到上新世現代各屬就已出現了。熊類以晚始新世至上新世還有兩個旁支。一個是犬熊類(Am-phicyoninae)。這類動物主要發現於歐洲大陸,北美也有少量化石。這是一類個體相當大的動物,其牙齒形態與犬科者很接近,所以長期來被歸入了犬科。近來發現它們頭骨基部構造和四肢的構造與熊的更為接近,才又改歸熊科。另一類是半熊類(Hemicyoninae),它們的地理分布與犬熊相同。早、中中新世以半熊(Hemicyon)為代表,晚中新世以印度熊(Indarctos)為代表,而上新世則以郊熊(Agriotherium)為代表。這也是一類個體很大的動物,牙齒與熊類者相當接近了。
鼬科(Mustelidae)現生種分布於除大洋洲、南極以及一些島嶼之外的各大洲。這是一些小至中型的動物。它們的裂齒都不大,裂葉沒有深的中央凹刻,上第一臼齒內半部加大,其長總比外緣長,第三臼齒退失,上、下第二臼齒也很退縮。鼬科在形態上分異很大,有很複雜的發育歷史。科以下有水獺類(Lutrinae)、鼬類(Mustelinae)、蜜獾類(Mellivorinae)、臭獾類(Mephitinae)、獾類(Melinae)、美獾類 (Leptaretinae)等亞科。鼬科動物的化石廣泛發現於全世界。最早發現於漸新世,中新世至現代十分繁盛。
犬科 (Canidae)現生種的地理分布大致與鼬科相同。它們大小中等,屬追捕型。頭骨低長,齒式較全,僅M3退失。它的進化主幹在北美,始於漸新世。在舊大陸從中新世末才開始出現,主要是貉(Nyctereutes)和犬(Canis),顯然是從北美通過白令陸橋遷入的。
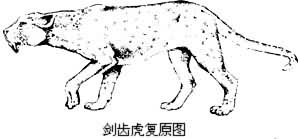
貓科 (Felidae)現生種分布與上相同。體型由小至大,屬躍捕型。頭骨短圓,裂齒與裂葉特別發達,犬齒粗大或十分扁長,其他頰齒退化。耳泡圓隆,分為二室,爪特別銳利,伸縮型。在早始新世末在北美出現了早期特化的獵貓類(Nimravinae)。它們的齒式較少退化,上裂齒保留有很大的原尖,但下裂齒下後尖消失。這類動物至晚中新世就絕滅了。歐洲大陸的後貓(Metailurus)有可能是它的後裔。另一支則是假劍齒虎類(Hoplophonei-nae)。它們的上犬齒已呈馬刀狀,下頜聯合部有下伸的葉狀突起。這類動物始於漸新世,分布於整個全世界。晚中新世出現了真劍齒虎(見圖),至更新世晚期才最後在北美滅絕。狹義的貓類大概是從歐洲漸新世的始貓(Proaelurus)經假貓(Pseudaelurus)逐漸演化而成。假貓和後期真貓化石在北美也有發現。
 食肉目(化石)
食肉目(化石)鬣狗科(Hyaenidae) 的現生代表是體型較大的追捕和食腐動物和一類特化的食蟻動物,僅分布於非洲和亞洲南部。食蟻的土狼(Proteles)牙齒已極退化,狹義的鬣狗則具粗壯的錐形前臼齒,裂齒髮育,臼齒退化。其進化主幹在古北界和非洲。中中新世時有一早期特化的中鬣狗類(Percrocuta)。晚中新世時鬣狗的種類很多,化石也很豐富,特別是其中的鼬鬣狗類(Ictitheriinae)。至上新世現生屬出現,其中有一支,豹鬣狗 ( Chasma-porthetes)通過白令陸橋進入北美。
