百科名片
頂復動物亞門(Apicomplexa)是動物界原生動物門的1門,為最原始、最簡單、最低等的單細胞動物。原生動物是單細胞,細胞內有特化的各種胞器。具有維持生命和延續後代所必需的一切功能,如行動、營養、呼吸、排泄和生殖等。每個原生動物都是一個完整的有機體。
 頂復動物亞門
頂復動物亞門簡介
原生動物分布廣泛。自由生活的種類能分布在海洋、陸地(包括淡水、鹽水、土壤、冰、雪以及溫泉中),甚至在空氣中也有原生動物的包囊;至於寄生的種類,則幾乎所有的多細胞動物都能被原生動物所寄生,植物也可成為原生動物的宿主;此外還有附生、共生、重寄生的類型.目前已描述的原生動物約6.8萬種,其中一半是化石種類,現生種類中,營自由生活的占2/3,寄生生活的占1/3。
多細胞動物的單個細胞一般不能脫離其他細胞而獨立生活,更不能象原生動物那樣具有生命的一切功能。原生動物包括相當多樣性的生物類群,在系統發育過程中它們可通過多種途徑進化到多細胞動物。例如海綿動物中就有與領鞭毛蟲十分相似的鞭毛細胞;群體的植物性鞭毛蟲的團藻蟲類開始有細胞間的功能分化——有專司營養和專司繁殖的個體,被認為在進化過程中可能是腔腸動物的祖先。人們推測無腸目的渦蟲可能起源於原始的多核纖毛蟲,從而構想低等的多細胞動物多發性地起源於原生動物的不同類群。
 頂復動物亞門
頂復動物亞門研究
300多年前,A.van列文虎克用約放大270倍的透鏡,第一個看到了很多自由生活和寄生生活的原生動物,他把眼蟲描述為“中間綠、兩端白”的蟲子,後人尊稱他為原生動物學之父。1758年C.von林奈套用雙名法命名了巨團藻蟲,大巨變蟲。G.A.戈爾德富斯1817年第一個用Protozoa(原生動物)一詞,但他把腔腸動物也包括進去。對原生動物下了正確定義的是C.T.E.von西博爾德(1845)。C.G.埃倫貝格1838年把觀察的原生動物進行分類描述,提出初步的分類系統。20世紀初,從原生動物的經典分類
 頂復動物亞門
頂復動物亞門研究逐步擴展到生理、營養、生態、遺傳等領域。50年代起,由於電子顯微鏡的套用,使原生動物的研究進入到亞顯微結構的水平。近十餘年來,套用顯微分光光度計、酶化學等方面的新技術,在分子水平上對原生動物進行微觀的研究。同時,通過套用計算機模型,也對原生動物的生態進行了群落級的巨觀研究。
形態
特徵
原生動物的形狀變化很大。有原生質隨意流動、形狀不定的變形蟲,有結構精巧、宛如雕刻著花紋的工藝品的放射蟲和有孔蟲。同一種類可因處於生活史的不同時期或不同的環境條件而改變形狀。個體大小一般在10微米~1毫米之間。最小的只有2微米(如寄生在紅細胞中的巴倍蟲,大的長達19厘米(如早新生代的錢幣蟲)。
細胞膜
原生質體外面有一層細胞膜,使原生動物和外界環境隔開。任何物質滲入體內或排出體外都必須通過這層細胞膜。植鞭毛綱中的衣滴蟲等在細胞膜外還包有細胞壁,但在眼蟲則為有彈性的表膜。正是由於這種表膜構造,使大多數原生動物既有比較穩定的形狀,又能夠行動自如。變形蟲的表膜薄,有利於伸出偽足。有些鞭毛蟲、肉足蟲、纖毛蟲有矽質、鈣質、纖維質的外殼,人們認為這些外殼的基質是在細胞內形成後再移到表膜外的。
細胞質
一般可分為透明、緻密的外質和液態、流動的內質。細胞質中含有各種顆粒(油滴、澱粉、副澱粉、色素等)和各種細胞器(線粒體、高爾基器、溶酶體等)。高爾基器在鞭毛蟲中很多,在纖毛蟲中缺如或不發達。高爾基器與細胞合成產物的精製、加工和儲存有關,如在合尾滴蟲、鱗殼蟲、放射蟲等體表呈圖案排列的鱗片,這些鱗片就是在高爾基器的泡囊內形成的,然後這些泡囊移到內質膜,鱗片再按一定的圖案排列在體表面。電鏡觀察揭示大多數纖毛蟲的線粒體的內膜向內突入,形成管狀的嵴,但在一些鞭毛蟲是片狀或盤狀嵴,在變形蟲和簇蟲是泡囊嵴。寄生在反芻動物胃中的厭氧纖毛蟲的線粒體是沒有嵴的囊,腸袋蟲的線粒體則是有嵴與無嵴的混合體。許多植物性鞭毛蟲和某些其他原生動物有呈紅-紫色的色素顆粒,稱為血色素,如雨紅球蟲、吉尼眼蟲、紅眼蟲、赭蟲。天藍喇叭蟲的外質含有1種喇叭色素,因而蟲呈藍色。
 頂復動物亞門
頂復動物亞門細胞核
根據染色質的構造可把細胞核分為泡狀核和緻密核兩類(圖1)。泡狀核常見於肉足蟲和鞭毛蟲,緻密核常見於纖毛蟲的大核中。在鞭毛蟲、肉足蟲、孢子蟲中,有的種類有很多細胞核,但都是同型核。許多原生動物在營養期間,細胞核的染色體是多倍體,有些則為二倍體或單倍體。纖毛蟲有大、小核之分,小核是二倍體,大核是高度多倍體。大核的大小、形狀、數量變化很大,在形狀上也各不相同差異很大。大、小核內均含脫氧核糖核酸(DNA)。一般地說小核控制細胞的遺傳,大核控制細胞的營養,但這不是絕對的。
已知大核在遺傳上能控制表型(見基因),說明大核中也有某些控制遺傳的因子。只有大核而沒有小核的品系雖然能分裂和生存,但最後還是會老化。在接合生殖過程中,大核退化,由小核產生新的大核,這說明大核雖有自主性,但仍要依賴小核。纖毛蟲的核二型性在整個生物界是獨特的。鞭毛蟲的細胞核與毛基體系統有密切關係。鞭毛的基體有根絲體聯繫到核上。基體與核在數量上有一定比例。在低等的鞭毛蟲中一般為2∶1,即2個基體,1個細胞核。隨著進化而擴大比例,在超鞭毛目中已是100~1000∶1了。
色素體及其附屬胞器
大部分植物性鞭毛蟲具有與光合作用有關的色素體和紅色的眼點。眼點由1至多個紅色小球構成,對光十分敏感,能引導鞭毛蟲游向陽光處。色素體有4類:葉綠素、胡蘿蔔素、葉黃素和藻膽素。由於所含色素體的組成和分量不同,植物性鞭毛蟲在顏色上差異很大,有綠、黃、藍等。
有色鞭毛蟲如長期處在黑暗而有機質豐富的環境中,色素體和眼點都退色,不再進行光合作用,它們用身體表面的滲透功能吸收營養或直接通過胞口吞入食物(如眼蟲)。
 頂復動物亞門
頂復動物亞門運動胞器
與鞭毛蟲、肉足蟲、纖毛蟲相應的運動胞器有鞭毛、偽足和纖毛。孢子蟲是寄生的,無專門的運動胞器,借身體的屈曲、滑動等方式移動,僅在生活史中的變形期,其小配子可借偽足和鞭毛運動。鞭毛蟲中除超鞭毛蟲有許多鞭毛外,一般常為1~2根,少數有8根。鞭毛是細胞質的絲狀突起,通常自身體前端伸出,少數自體側、腰溝、體後伸出成舵鞭毛。鞭毛還有捕食、附著、感覺的功能。它由兩部分組成:中央為1根有彈性的、由若干平行的纖維組成的軸絲,軸絲外部包著原生質鞘(圖2)。原生動物中的鞭毛和一切真核生物中的鞭毛一樣,是“9+2”的格式(見鞭毛與纖毛)。
鞭毛是從表膜下埋在細胞質內的、可染色的毛基體亦稱基體,在纖毛蟲中稱為動基體延伸到體外。一般鞭毛蟲有2個基體,位在細胞頂端,但只有一個基體上伸出一根鞭毛,另一個基體是禿的(如低等的錐蟲)或有一點鞭毛的痕跡(如眼蟲)。在較高等的動鞭毛蟲中,基體可有2、6、8甚至數百個不等。在超鞭毛蟲中有2000根左右的鞭毛,基體數量總比鞭毛多5個,這5個不長鞭毛的基體在個體發育上具有重要的意義。基體本身不分裂,新的基體在靠近原有的基體處形成。電子顯微鏡觀察證實基體的外周微管排列格式與鞭毛相似,只是在雙重體中增加了第3根,成為三重體(圖3f、g)。在寄生的鞭毛蟲中與鞭毛有關的構造還有波動膜、肋、脊突、副基器、副基絲、軸桿、盾、錘、紡錘簇等。鞭毛自頂端向基部作波浪運動,有平面的和螺鏇的兩種波浪運動形式,均可使蟲體向任何方向移動。偽足根據形態可分為葉足、絲足、根足(又稱網足)和軸足四類。偽足除了行動之外,還有捕食、固著、感覺的功能。關於偽足的變形運動已研究了100多年,至今仍未解決。一般認為當變形蟲行動時,內質從半液態的溶膠轉化為半固態的凝膠,凝膠有收縮性,它對溶膠產生微壓,於是溶膠便向最薄弱的地方流去,並形成偽足。因此偽足象個凝膠管,溶膠被迫向管子流去。
 頂復動物亞門
頂復動物亞門溶膠達到凝膠管前端接近表膜處就轉化為凝膠。在後部收縮的凝膠又轉化為新的溶膠(圖4)。如此不斷重複,溶膠不斷向前流動,身體後部就不斷縮小,結果偽足便增大而成為身體的主要部分,同時前面又不斷形成新的偽足。這種變形運動常見於葉足型的種類。絲足、網足、軸足的運動便是另一形式,因為這類偽足很細,中央有較硬的纖維,溶膠象傳送帶似地沿著纖維向兩個相反方向流動。纖毛與鞭毛一樣,也是由軸絲和鞘組成,軸絲的亞顯微結構也是按“9+2”的格式排列(圖5)。
每根纖毛的基體伸出一細纖維,位於基體的左邊,並與同一行列內其他纖毛基體伸出的纖絲相連結,成為縱行的動纖絲。纖毛比鞭毛短而數量多。纖毛有運動、捕食和感覺的功能,而且隨分工不同而特化。有的纖毛癒合成小膜,在口緣成帶狀排列,動作有力而協調,便於將食物驅入口中。有的纖毛癒合成大的波動膜,突出在口緣之外。在下毛類中,背部纖毛退化,腹部纖毛融合成毛筆狀的棘毛。原生動物中運動最快的是纖毛蟲。每根纖毛在打水時,分效力擊打和恢復擊打兩種。整個纖毛的協調擊打、沿著身體產生4種類型的節律波。
纖維構造
大多數原生動物都有收縮性,有的種類特別明顯,如喇叭蟲和鏇口蟲的肌絲,鍾蟲柄內的牽縮絲,其收縮性都很強。電子顯微鏡證明細胞質內都有纖維結構,除了能幫助身體收縮外,還有幫助運動(如簇蟲亞綱)、保持體形(如蛙片蟲、小瓜蟲等)的功能。
支持和保護胞器
這類胞器是細胞外的構造,如柄、殼、內外骨骼、孢囊、孢子等,這些結構是多細胞動物所沒有的。屬於內骨骼的如動鞭毛蟲的軸桿、肋,放射蟲輻射伸出的刺或骨針,裸口類纖毛蟲口器內的咽籃、刺桿等,多半是起支持和保護的作用。屬於外骨骼最常見的是在加厚的表膜外有一層保護的殼。殼有幾丁質、矽質、硫酸鍶等成分,有時還有石灰質沉澱。
有的殼是整片的,有的是鱗片狀的,有的有精美雕刻的圖案。原生動物還能從自體內射出各種突出質,用以進行防衛、攻擊和取食。纖毛蟲有7種突出質:刺絲泡(如草履蟲)、粘絲泡(如四膜蟲)、毒絲泡(如長頸蟲)、系絲泡(在吸管蟲的觸手上)、纖絲泡(如擬小胸蟲)、桿絲泡(如管刺蟲)和網絲泡(如櫛毛蟲)。鞭毛蟲中也有刺絲泡和粘絲泡,如腰鞭毛蟲有刺絲囊、粘孢子蟲有極絲,它們起著固著的作用。
伸縮泡和其他各種液泡
生活在淡水的原生動物有1至多個伸縮泡生活在海水中的和寄生的原生動物一般沒有伸縮泡。變形蟲只有1個伸縮泡,構造簡單,位置不定,伸縮泡的周圍有小囊泡,線粒體密度較大(圖6)。草履蟲有前後兩個伸縮泡,各有1箇中央泡和6根輻射伸出的收集管,收集管上有分枝小管與內質網的管道系統相連(圖6)。
伸縮泡的主要功能是調節滲透壓。食物泡恨據攝食情況分為兩種:含大顆粒食物的吞噬泡和含溶解性營養物質的吞飲泡。食物顆粒或營養物質進入食物泡後,食物泡與細胞質內的溶酶體合併,食物被溶酶體釋放的酶消化,未經消化的廢物被排出體外。
繁殖和生命周期
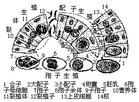
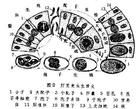
原生動物的生命周期包括生殖期和孢囊。有些種類已失去形成孢囊的能力。生殖期可分為無性生殖和有性生殖。大多數原生動物無性生殖用二分裂法。鞭毛蟲是縱分裂,纖毛蟲是橫分裂。緣毛類纖毛蟲外表看來如縱分裂,但是細胞內各成分(如核、口圍纖毛帶)仍是橫分裂。瘧原蟲、球蟲則行裂體生殖(復分裂)。吸管蟲在體內或體外生出許多芽體(出芽)。
有些多核的原生動物如膠絲蟲、多核變形蟲偶爾會分裂成2至數個小的、仍是多核的個體(原生質團分割)。以上4種方式只有出芽生殖還保留親體,其餘均無親體,後代都是等同的。原生動物有性生殖有3種:融合、接合、自體受精和假配。自體受精與接合生殖相似,但只在一個個體內進行。小核分裂數次,其中有兩個配子核融合成合核,其餘退化。合核分裂形成新的大、小核。假配與接合生殖一樣,要求兩個個體接觸,但沒有配子核的交換,每個個體完成自體受精過程後各自分開。在草履蟲中曾發現過這兩種特殊的核現象。
寄生原生動物的生活史比較複雜。大多數孢子生活史包括3個時期:裂體生殖期、配子生殖期和孢子生殖期。有明顯的無性世代與有性世代的交替。孢子生殖期是由合子產生的孢子母細胞形成孢子後,再進一步形成子孢子。子孢子一般包有外殼,能抵抗不良環境,有利於傳布。
生態
影響原生動物的環境因子有以下幾個方面:
①水分,原生動物不論怎樣小也要求有最低限度的水分。
②溫度,原生動物可生活在地球的兩極和冰雪之中,也可生活在溫度較高的溫泉中,如中國的西藏自治區67℃的溫泉中,就生活著鰩頜砂殼蟲。在實驗室內可用冰凍方法保存原生動物,在-95℃時梨形四膜蟲能生存4個月,寄生的牛胚毛滴蟲能生存5年半。天然條件下5~35℃對大多數原生動物是適宜的。
③溶解氧,大部分原生動物需要氧氣以維持生命活動。但在深水湖的湖底、污水厭氧發酵池中生活的鞭毛蟲、纖毛蟲是厭氧的,大多數寄生原生動物也是厭氧的。
④溶解的二氧化碳,大多數原生動物能耐受低濃度的CO2。對植物性鞭毛蟲來說,CO2是重要的碳源。
⑤PH,在PH值是2.2~9.2的範圍內都可能生存。
⑥鹽度,原生動物能分布在淡水、鹹淡水、海水、鹽水中。大多數種類只能生存在一定的鹽度範圍內,有的種類是廣鹽性的,利用伸縮泡調節體內的滲透壓以適應環境鹽度的變化。
⑦光,光是植物性鞭毛蟲進行光合作用的能量來源。在強光下,團藻蟲和眼蟲能躲避。在西藏雪山上看到的衣滴蟲能產生抵抗強光的保護色素——紅色素,致使積雪呈紅色。
⑧底質、水流、風浪等,放射蟲身體中的空泡能擴大和縮小,以調節自身在水中的深度,不致受風浪的吹逐。有一種生活在潮間帶水潭內的眼急游蟲,其生活習性能符合潮汐的節奏。退潮時水潭露出,眼急游蟲游到水面取食、分裂和繁殖。在潮水來到之前,它就到水潭的底部形成包囊。包囊粘於底部,以免被潮水帶走。
⑨食物和營養,營養方式有植物式、動物式和腐生式3種。大部分寄生原生動物是腐生式營養,近來發現有幾種孢子蟲能用一種特別的胞口吞食宿主紅細胞的細胞質而形成食物泡。此外,原生動物還需要某些生長因子如維生素B1、B12、H等。寄生原生動物是從自由生活的原生動物演化來的,在長期適應過程中形態和生理髮生了變化,如失去了攝食胞器,增設了固著胞器(吸盤、極絲、柄等),提高了繁殖能力和完善了轉移新宿主的途徑(從單一宿主發展為中間宿主和儲存宿主)。
此外,原生動物也可被其他生物寄生或與其他生物共生,例如,有孔蟲中有寄生的小變形蟲,簇蟲體內有寄生的微孢子蟲,雙小核草履蟲體內的Kappa粒也是細菌,在許多纖毛蟲體內都發現有共生綠藻。
變異與遺傳
基因變異的潛力在雙倍體和多倍體的原生動物種群中比單倍體的大,在有性生殖過程中又比無性生殖過程中大。在衣滴蟲的不同性和纖毛蟲中不同交配型之間進行交叉受精,就能擴大新基因組合的範圍。纖毛蟲進行接合生殖必須具備兩個條件:
①兩個體必須屬於適當的交配型。
②其交配型純系必須是成熟的齡期。在一個種內有數個遺傳上相分離的基因群,每個基因群有相同的基因庫,但有幾個不同的交配型。在同一基因群內的交配型互相接合的幾率可達95%。已知雙小核草履蟲有14個基因群,各有兩個交配型,綠草履蟲有6個基因群,各基因群內有2~3個交配型。在原生動物中下毛目是進化比較高等的一類,其皮層上有許多按一定格局分布的複雜的纖毛和非纖毛結構。
這些結構的遺傳是自主性的。纖毛蟲纖毛模式的形成和決定很難完全歸結為基因對表型的控制作用或是某個基因產物的作用。在皮層和毛基體內至今還沒有找到遺傳信息的載體DNA分子。因此有人認為遺傳性不是核酸分子所能獨攬的。
系統發育
一般認為原生動物的祖先是一些古老的“植物-動物”性的類群,稱為古代的植鞭毛蟲。從古代植鞭毛蟲分為幾個不同的根株,每個根株上升到現代原生動物的祖先。這是一種多系統的理論。有人認為從古代植鞭毛蟲演化為肉足蟲。孢子蟲是雙重起源的,一類起源於肉足蟲,一類起源於動鞭毛蟲。纖毛蟲直接起源於鞭毛蟲,只是還說不清楚大、小核的分化和接合生殖的演化。
無論哪種觀點都是從現代的原生動物中進行推測,缺乏充分的根據。經典的分類把原生動物門分為4個綱——鞭毛蟲綱、肉足蟲綱、孢子蟲綱和纖毛蟲綱。隨著發現種類的增加,新技術(如電子顯微鏡等)的套用,分類學家對原生動物傳統的分類系統進行了修正。比較一致的有。
①鑒於許多鞭毛蟲的生活史中有變形期,許多肉足蟲的生活史中有鞭毛期,有的種類本身就兼有鞭毛和偽足,所以把兩大綱合併為肉鞭動物亞門。
②傳統的孢子蟲綱內有些種類的生活史中並不出現孢子,應當分出來。電子顯微鏡觀察發現某些種類在子孢子或裂殖子時期其頂端有一個複雜的亞顯微結構——由極環、類椎體、表膜下微管、微孔、棒狀體、微絲組成的頂復體,因而將其獨立為頂復動物亞門,與微孢子蟲亞門、粘體動物亞門、囊孢子蟲亞門並列。
③傳統的分類中把盤蜷蟲類放在肉足綱內。現已證明它的絲網並不是偽足,而是堅硬的、無生命的絲,因而獨立為盤蜷動物亞門。
分類
下列的分類系統主要參照原生動物學家協會分類和進化委員會主席N.D.萊文於1980年與16位分類學家協商的一個方案。本文把該方案中7個門下降為7個亞門、亞門下降為超綱,在肉足超綱內設有綱、亞綱和總目,原生動物仍作為一個門:
原生動物門(Protozoa)
肉鞭動物亞門(Sarcomastigophora)
鞭毛總綱(Mastigophora)
植鞭毛綱(Phytomastigophorea)
隱滴蟲目(Cryptomonadida)
腰鞭目(Dinoflagellida)
眼蟲目(Euglenida)
頂復動物---瘧原菌
金滴蟲目(Chrysomonadida)
異鞭目(Heterochlorida)
綠滴蟲目(Chloromonadida)
定鞭目(Prymnesiida)
團藻蟲目(Volvocida)
溪滴蟲目(Prasinomonadida)
矽鞭目(Silicoflagellida)
動鞭毛綱(Zoomastigophorea)
頜鞭目(Choanoflagellida)
動體目(Kinetoplastida)
原滴蟲目(Proteromonadida)
曲滴蟲目(Retortamonadida)
雙滴蟲目(Diplomonadida)
銳滴蟲目(Oxymonadida)
毛滴蟲目(Trichomonadida)
超鞭自(Hypermastigida)
蛙片總綱(Opalinata)
蛙片綱(Opalinatea)
蛙片目(Opalinida)
肉足總綱(Sarcodida)
根足綱(Rhizopodea)
葉足亞綱(Lobosia)
裸變總目(Gymnamoebidea)
變形目(Amoebida)
裂芡目(Schizopyrenida)
生目(Pelobiontida)
殼葉總目(Testacealobosidea)
表殼目(Arcellinida)
毛片目(Trichosida)
微膠絲亞綱(Acarpomyxia)
細膠絲目(Leptomyxida)
堅膠絲目(Stereomyxida)
混膠絲亞綱(Acrasia)
混膠絲目(Acrasida)
真膠絲亞綱(Eumycetozoia)
原星總目(Protosteliidea)
原星目(Protosteliida)
網星總目(Dictyosteliidea)
網星目(Dictyosteliida)
膠胚總目(Myxogastridea)
