
主要特徵
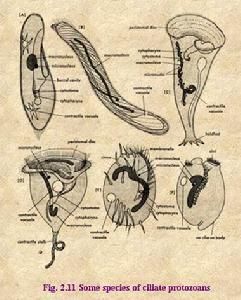 纖毛綱
纖毛綱纖毛蟲分游泳型和固著型兩種類型,他們以纖毛作為運動和攝食的細胞器。纖毛蟲是原生動物中最高級的一類,它們有固著的、結構細緻的攝食細胞器。固著型纖毛蟲大多數有肌原纖維,細胞核有大核(營養核)和小核(生殖核)。纖毛的結構與鞭毛相同,其不同點是纖毛較短,數目較多,運動時節律性強。纖毛可成排分散存在,也可由多數纖毛粘合成小膜、排列在口的邊緣稱為小膜帶。也可由一單排纖毛粘合形成波動膜,通常在胞咽中。還有的纖毛成簇粘合成束稱為棘毛(在蟲體腹面爬行用)。
結構一般較複雜。原生動物中這類動物是分化最多的。細胞核一般分化出大核與小核。大部分纖毛蟲具有攝自的胞器。
形態與結構
纖毛蟲類是分布極廣泛的原生動物,任何水域,甚至污水溝也有分布。大多數為單體自由生活,少數群體營固著生活。也有少數營共生或寄生生活。體長一般在10μm—3mm之間。體形變化很大,一般遊動的種類呈長圓形或卵圓形,爬行生活的種類多呈扁平形。固著生活的種類具有長柄,還有極少數種類可以分泌粘液或用外來物質粘合成兜甲。體表均有表膜,表膜外全身或部分披有纖毛或纖毛的衍生物,內質與外質分化明顯,外質中還含有複雜的結構,內質中有較多的顆粒,含有食物泡、伸縮泡、細胞核等結構。最熟悉的纖毛蟲綱代表種有草履蟲(Parameciumcaudatum)及四膜蟲(Tetrahymena)。纖毛蟲類的最外表有一層表膜,有的種類是一層很薄的薄膜,草覆蟲及許多高等的種類表膜呈現整齊排列的突起及凹陷,在光學顯微鏡下表膜表現成無數整齊排列的六角形小區。小區的中央即為凹陷部分,稱纖毛囊(ciliarycapsule),由此伸出1—2根纖毛。纖毛也整齊排列。突出部分是由於纖毛基部附近形成的表膜小泡(alveolus),以致成突起狀,它的存在增加了表膜的硬度,固定了纖毛及刺絲泡的位置,有利於體形的維持。纖毛與鞭毛的結構相似,也是由典型的“9+2”微管纖維組成,基體位於外質中纖毛的基部,但纖毛的基體在其一側發出1—2條很細小的纖維,稱細動纖絲(kinetodesmalfibril),它前行一段距離之後,與同行其他基體發出的細動纖絲聯合,形成一較粗的縱行的動纖絲(kinetodesmata),構成了纖毛蟲的下纖列系統(infraciliature)。由基體還向深層發出縱行絲,其中含有肌球蛋白及肌動蛋白,這說明纖毛運動的性質與肌肉一樣,也是由肌動蛋白絲在肌球蛋白絲上滑動所引起。也有人認為下纖列系統具有神經傳導及協調纖毛運動的作用,因為他們發現纖毛蟲類也具有乙醯膽鹼及乙醯膽鹼酯酶。也有人認為纖毛的協調運動是由於細胞膜的去極化作用,與神經衝動的傳遞作用相似。下纖列系統為纖毛蟲類所特有,在一些成蟲期纖毛消失的種類如吸管蟲(Suctorida)卻仍保留有下纖列系統。在外質的深部與纖毛相間排列的是一些棒狀或卵圓形小體,垂直於體表,成行排列,稱為刺絲泡(trichocyst)。它們或散布整個體表,或限制在體表的一定區域。刺絲泡未排放時呈管狀,或囊狀,當遇到各種理化刺激時,囊中的物質排出,由於吸水聚合而成為刺絲,刺絲通常有毒,用以捕獲及麻醉其他動物,或用於自身防衛。纖毛蟲類的纖毛由於著生部位及功用的不同,可以分為體纖毛及口纖毛。體纖毛著生在身體表面,在原始的纖毛蟲,纖毛是均勻的覆蓋在整個身體表面,較高等的種類纖毛僅分布在身體的一定區域,或纖毛癒合成棘毛(cirrus)。體纖毛的作用是運動。口纖毛著生在口區,常成排的纖毛粘著成板狀,或再由許多纖毛板聯合成帶狀,稱為小膜帶(membranella),或是更多的纖毛單行排列成波動膜(undulatingmembra-ne),由於小膜帶或波動膜上纖毛的協調擺動,用以收集或傳送食物。[1]
常見種類
纖毛蟲的種類很多。不同類的纖毛蟲,其纖毛的多少和分布的位置是不同的。有些全身改有纖毛(屬全毛類),如草履蟲、小瓜蟲等。小瓜蟲(Ichthyophthirius)寄生在魚的皮膚下層、鰓、鰭等處。這種纖毛蟲對魚危害很大,病魚的死亡率很高。用百萬分之一的硝酸亞汞給魚洗澡效果較好有些種類纖毛不發達,僅限於蟲體的腹面(屬下毛類),如棘尾蟲(Stylonychia)、游仆蟲(Euplotes),用腹面粗大的棘毛爬行。有些是纖毛在圍口部形成口緣小膜帶(屬緣毛類),如鍾蟲(Vorticella),口緣小膜帶由左向右旋,其他部分無纖毛,體下端有一能伸縮的棲,可營固著生話。又如車輪蟲(Trichodina),寄生於淡水魚的鰓或體表。
自由生活的纖毛蟲,大部分為浮游生物的組成部分,是魚類的餌料。
代表生物
草履蟲
也是原生動物的代表種。世界已知22種,常見有大草履蟲(長180~300微米)、雙小核草履蟲(長80~170微米,伸縮泡2個,有兩個小核,很小)、多小核草履蟲(長180~310微米,有時有3個伸縮泡,小核泡型,有3~12個)、綠草履蟲(體長80~150微米,細胞質內有綠藻共生,在見光處培養後通體呈綠色,小核1個,緻密型)。大草履蟲Parameciumcaudatum是最常見者,屬膜口目草履蟲科大草履蟲屬。生活在有機質較豐富的池塘、緩流的小溝、小河以及居民區附近的水溝中。大草履蟲是原生動物中體形較大的種類,體長約180~300微米,用肉眼觀察含有大量草履蟲的液體,可以看到很多針尖狀發亮的白色小點在浮動,這些白點就是草履蟲。草履蟲一般呈長圓筒形,前端鈍圓,後端寬而略尖,形狀似倒置的草鞋,因此得名。草履蟲全身長滿縱行排列的纖毛,纖毛和鞭毛的結構基本相同。蟲體的表面為表膜,由3層膜組成,起到緩衝和保護作用,裡面的細胞質分化為內質與外質。每一根纖毛從位於表膜下的一個基體發出來,整個表膜下的基體由縱橫連線的小纖維連線成網,起到協調纖毛活動的作用。表膜下的外質中有一排小桿狀的囊泡結構,與表膜垂直排列,叫做刺絲泡,開口於表膜上,當蟲體遇到刺激時,刺絲泡射出其內容物,遇水形成細絲,一般認為具有防衛和捕食的作用。草履蟲營吞噬營養,有較複雜的消化細胞器。由蟲體近前端開始有一條口溝,斜著伸向身體中部,口溝末端的與表膜相連處形成胞口,下面連著漏斗形的胞咽。食物(如細菌、小的生物和腐爛的有機物等)通過口溝處纖毛擺動而進入胞口,在胞咽下端積聚成下泡,被細胞質包裹而脹大,形成食物泡,在固定的路徑中流動,與溶酶體融合後進行消化。最後不能消化的殘渣由身體後部的胞肛排出蟲體外。在內、外質之間有2個伸縮泡,一個在體前部,一個在體後部。每個伸縮泡向周圍細胞質伸出放射排列的收集管,這些收集管端部與內質網的小管相通。在伸縮泡主泡和收集管上有由一束微管組成的收縮絲,內質網收集的水分以及代謝廢物通過收縮絲的收縮而進入收集管,注入伸縮泡,從表膜小孔(排泄孔)排出蟲體外。前後2個伸縮泡交替收縮,維持其體內水分平衡。可見,伸縮泡的功能是調節滲透壓和排泄。草履蟲的呼吸作用主要通過體表進行,由體表吸取水中的氧氣,將新陳代謝產生的二氧化碳排出體外。大草履蟲有2個細胞核,大核與營養有關,小核與遺傳有關。生殖可分為無性生殖和有性生殖。無性生殖為橫二分裂,小核先進行有絲分裂,大核再進行無絲分裂,然後細胞質一分為二,最後蟲體從中部橫斷,成為2個新個體。有性生殖為接合生殖,2個草履蟲口溝對口溝粘合在一起,粘合處細胞膜溶解,兩個蟲體的細胞質相通,小核離開大核,分裂2次形成4個核,其中3個解體,剩下的1個小核又分裂成大小不等的2個核,大核逐漸消失。然後兩蟲體互換其新小核,並與對方較大核融合,這一過程相當於受精作用。此後兩蟲體分開,接合核分裂3次成為8個核,4個變為大核,其餘4核有3個解體,剩下1核分裂為2小核,再分裂為4小核,每個蟲體也分裂2次,結果原接合的兩親本蟲體各形成4個草履蟲,新形成的8個草履蟲都有一大核,一小核。
鍾蟲
體呈吊鐘形,鐘口盤狀口區周圍有一腫脹的鑲邊,其內緣著生三圈反時針旋轉的纖毛(他處概無纖毛)。口盤與鑲邊均能向內收縮。口自鑲邊內緣斜入體內,有一振動的波動膜。大核馬蹄形,小核粒狀。身體反口面的頂端有一長柄,用以附著他物,內有肌束,當蟲體收縮時,也可螺旋狀捲曲。產於淡水中。單體,但常簇生。我國已發現111種。鍾狀身體的底部收縮為帚胚,由此長出能伸縮的、內含肌絲的柄,以固著在各種基質上。成體營固著生活,體纖毛退化。前連線埠圍部分的纖毛十分發達,由3片左旋小膜口圍帶組成。以細菌、碎屑或藻類為食。無性生殖。群體生活的種類,柄分叉呈樹枝狀、每根枝的末端掛了鐘形的蟲體。無論是單個的或是群體的種類,在廢水生物處理廠的曝氣池和濾池中生長十分豐富,能促進活性污泥的凝絮作用,並能大量捕食游離細菌而使出水澄清。因此,它們是監測廢處理水效果和預報出水質量的指示生物。
喇叭蟲
體形似喇叭,表面披有均勻的纖毛,後端尖細,呈柄狀;前端似喇叭口,周緣有許多纖毛癒合成的膜狀毛,口即位於近旁,為動物性營養。體內有念珠狀的大核(與營養有關)、若干小核(與生殖有關)與伸縮泡。生活於池沼中,常以柄附著他物上,也能自由游泳。一般體極微小,須用顯微鏡觀察,較大的種,則肉眼可見。
車輪蟲
蟲體側面觀如氈帽狀,反面觀圓碟形,運動是如車輪轉動樣。隆起的一面為前面或稱口面,相對而凹入的一面為反口面。口面上有向左或反時針發方向螺旋狀環繞的口溝,其末端通向胞口。口溝兩側個生一行纖毛,形成口帶,直達前庭腔。反口面的中間為齒環和輻線環。在輻線環上方有一馬蹄形的大核,一個長形的小核和一個伸縮泡,其中部想體內凹入,形成附著盤,用於吸附在宿主身上。車輪蟲用附著盤附著在魚體的鰓絲或皮膚上,並來回滑動。游泳時一般用反口面向前像車輪一樣轉動,所以稱為車輪蟲。生殖用縱二分裂法和接合生殖。[1]
詞條圖冊
