
名稱來源
病毒粒子呈周期性間隔的異常整齊排列,形成晶格平面並互相重疊,當有斜射光線照射時呈現藍色或紫色虹彩,故名虹彩病毒(Iridos希臘字,意指希臘神話中為諸神報信的虹彩女神)。
比較複雜
最初多從昆蟲體中分離獲得,但是以後陸續地從許多其它動物如爬行動物、魚類、軟體動物、原蟲、乃至藻類和植物體內分離到了虹彩病毒。其中多數虹彩病毒是非致病性的,但有些虹彩病毒可引起魚類、蛙等動物的疾病,危害比較大,也有些虹彩病毒被人類用來在農業生產上防治病蟲害。如二化螟虹彩病毒(CⅣ)可以感染黑尾葉蟬〖WT5BX〗(Nephtotottix cincticeps),並可使深山葉蟬(Colladonus montanus) 發生致死感染,而後者正是傳播核果支原體病的傳染媒介,從而可防治本病。近幾年來對虹彩病毒的研究逐漸增多,不僅從各類動物、植物中分離出了多種屬於虹彩病毒科的病毒,而且對虹彩病毒的形態結構特點,特別是虹彩病毒的分子生物學特性進行了細緻的研究,同時在虹彩病毒的分類學方面也取得了很大的進展。根據國際病毒分類委員會〖WT5BZ〗(ICTV)的第六次報告,虹彩病毒科〖WT5BX〗(Iridoviridae)下設五個屬:即虹彩病毒屬(Iridovirus),綠虹彩病毒屬(Chloriridovirus),蛙病毒屬(Ranavirus),淋巴囊腫病毒屬(Lymphocystivirus)和“類金魚1型病毒屬”〖WT5BZ〗(“Goldfish virus 1?like viruses”)。非洲豬瘟病毒原來也屬於虹彩病毒科,後因發現其DNA結構和複製方式明顯不同於其他虹彩病毒,故已由虹彩病毒科中分離出去,列為未分類的病毒。虹彩病毒的病毒粒子較大,病毒粒子直徑為125~300nm。結構比較複雜,它具有緻密的不規則的中央核心體,即含有雙鏈DNA的病毒核心體。昆蟲虹彩病毒沒有囊膜,脊椎動物的虹彩病毒有囊膜,但同一樣品的病毒粒子可具有一或兩層環繞的膜。病毒的球狀核蛋白核心被富含脂質的結構蛋白亞單位組成的膜所包圍,某些病毒的病毒粒子成熟後再獲得一個來自胞漿膜的囊膜。該囊膜可以增強病毒的感染性,但它對病毒增殖並不是必需的。囊膜內為一個二十面立體對稱的衣殼,衣殼由多達1 892或2 172個殼粒組成。虹彩病毒粒子的分子量為500~2 000×10?6 Da,S??20W?=1 300~4 450,CsCl?2中的浮密度為1?35g~1?60g/cm?3。虹彩病毒屬和綠虹彩病毒屬的成員因沒有含脂質的囊膜而對乙醚有抵抗力,而其它屬病毒有囊膜對乙醚和非離子去污劑敏感。本科病毒在pH3~10條件下穩定,在4℃條件下可存活數年。對熱敏感,55℃15~30分鐘即可使其滅活。〖KH4〗〖HT5”〗〖JZ〗圖34-1〓虹彩病毒的核衣殼??〖HT5SS〗。
結構
虹彩病毒的核衣殼內含有單分子的線狀雙股DNA,長為170~200 kb,其分子量為100~250×10?6 Da,帶喙伊蚊虹彩病毒的基因組有440 kb。有些病毒可能在衣殼內具有雙分子核酸。核酸占整個病毒粒子總重量的12%~30%,G+C含量為29%~32%,但蛙病毒屬高達58%。虹彩病毒的核酸有其獨特的結構:在雙股DNA分子末端都具有單鏈結構;在蛙病毒屬、淋巴囊腫病毒屬及虹彩病毒屬(至少一種病毒)的基因組雙鏈DNA分子鏈兩端都帶有同一基因序列,,稱之謂“末端過剩”(terminanl redundancy),而且在不同分子的兩端又呈現不同的末端環狀排列,稱之謂“環狀變換”(circular permutation),這是虹彩病毒基因組的一個基本共同特點。但是在虹彩病毒屬和蛙病毒屬的代表株之間以及蛙病毒屬和淋巴囊腫病毒屬的代表株之間沒有發現基因組DNA的同源序列。虹彩病毒的基因組DNA另一個特點是胞密啶5′端的高度甲基化,特別是蛙病毒和淋巴囊腫病毒(因為二者的基因所編碼DNA甲基轉移酶的緣故)。甲基化可能與保護DNA免受酶切和與DNA複製、重組、修復及其基因表達有關。虹彩病毒具有20種以上的結構蛋白和若干病毒粒子結合性酶。以單向和雙向PAGE技術研究病毒結構蛋白發現有13~35種結構多肽,分子量在10~250×10 Da範圍內。
酶活性
病毒粒子中具有酶活性,特別是蛋白激酶活性。目前已研究證明二化螟虹彩病毒有RNA聚合酶、蛋白激酶、核苷三磷酸水解酶以及鹼性蛋白酶等活性,後者可能是一種絲氨酸蛋白酶,在蛙病毒3型病毒粒子中至少有6種酶活性,除蛋白激酶外,在核衣殼內還有核苷三磷酸水解酶、胸腺嘧啶激酶和DNA甲基轉移酶等。無囊膜的虹彩病毒(如昆蟲虹彩病毒)的病毒粒子中含有5%~9%的類脂質(主要是磷脂),它是構成正二十面體衣殼整體不可缺少的成分。有囊膜病毒(如從蛙、魚分離的虹彩病毒)在其來自宿主胞漿膜的囊膜中也含有類脂質,約3%~14%,其它成員類脂質的含量取決於囊膜的形成方式。在虹彩病毒的粒子中未檢出有糖類存在。在病毒的抗原方面,不同昆蟲的虹彩病毒之間在血清學反應上具有廣泛的交叉反應,但是昆蟲虹彩病毒與脊椎動物虹彩病毒之間無交叉反應,各種脊椎動物的虹彩病毒之間也無交叉反應。實驗證明,針對虹彩病毒的抗體一般對病毒粒子無中和作用。目前尚未對虹彩病毒進行系統的血清學調查,但已有的研究表明脊椎動物虹彩病毒和非脊椎動物虹彩病毒之間在血清學和核酸序列上均無關。
胞飲作用
虹彩病毒通過胞飲作用進入宿主細胞,並在吞飲液內脫衣殼。病毒感染早期即迅速抑制宿主的大分子合成,病毒的轉錄至少分為三期,即立即早期、早期和晚期,晚期轉錄並不依賴DNA的複製。但脊椎動物虹彩病毒缺乏合成mRNA必需的酶,故需依賴宿主細胞的RNA聚合酶Ⅱ。病毒DNA的複製可分為兩個階段:第一階段在感染早期,在宿主細胞核內合成較小的複製型DNA分子,其長度等於或比基因組小二倍;第二階段是在感染的晚期(在晚期基因表達後),在胞漿內複製成極大的複製複合體,即DNA連結體,並在胞漿內加工切割成成熟的DNA,包裝入成熟的病毒顆粒。這種加工結果可能導致“末端過剩”和“環狀變換”的形成。成熟病毒粒子的裝配發生在胞漿中,成熟病毒的釋放是通過胞漿膜上出芽或細胞溶解,出芽釋放的病毒獲得一個來自原生質膜或內質網膜的囊膜。
細胞漿中增殖
虹彩病毒可以在較廣泛的培養細胞上生長,並對感染細胞具有抑制其大分子生物合成的作用。虹彩病毒在細胞漿中的增殖與痘病毒相似,但虹彩病毒的核酸合成既依賴於宿主細胞漿,又依賴於宿主的細胞核。
位置
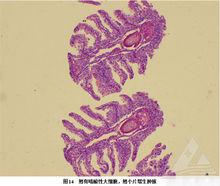 真鯛虹彩病毒病症圖
真鯛虹彩病毒病症圖病毒裝配是在細胞漿中進行的,在電子顯微鏡下觀察, 可見到胞漿中有大量堆集的二十面體立體對稱的虹彩病毒。昆蟲虹彩病毒的感染範圍限於無脊椎動物。易在來自其自然宿主的培養細胞上增殖,虹彩病毒32型病毒既可感染陸生等足目動物,又可感染線蟲。虹彩病毒僅能從其生活史中有水生階段的變溫動物體內分離出,大多數昆蟲虹彩病毒能使感染幼蟲變藍或青綠色,但脊椎動物虹彩病毒不能引起顏色變化。帶喙伊蚊虹彩病毒能通過孵巢垂直傳播,而一些虹彩病毒可水平傳播。
