流動鑲嵌模型(fluidmosaicmodel)
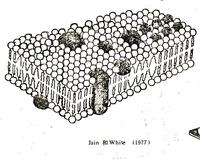 流動鑲嵌膜模型
流動鑲嵌膜模型流動相嵌模型有兩個主要特點。其一,蛋白質不是伸展的片層,而是以摺疊的球形鑲嵌在脂雙層中,蛋白質與膜脂的結合程度取決於膜蛋白中胺基酸的性質。第二個特點就是膜具有一定的流動性,不再是封閉的片狀結構,以適應細胞各種功能的需要。
這一模型強調了膜的流動由性和不對稱性,較好地體現細胞的功能特點,被廣泛接受,也得到許多實驗的支持。後來又發現碳水化合物是以糖脂或糖蛋白的形式存在於膜的外側表面。
【細胞膜的流動性和不對稱性】
膜的流動性是細胞膜結構的基本特徵之一,同時也是細胞膜表現其正常功能的必要條件。膜的流動性是指膜結構分子的運動性,它包括膜脂的運動和膜蛋白的運動。多年來,人們藉助於不斷創新的實驗技術,對於膜結構特性的研究進一步加深,並不斷取得進展。
(1)膜脂的流動性
在正常生理狀況下,膜脂分子處於運動狀態。膜脂的運動方式主要有側向擴散、鏇轉運動、鏇轉異構運動、左右擺動以及翻轉運動等。膜脂的流動性受著一些因素的影響,主要影響因素有:①溫度:在一定溫度下,脂分子從液晶態(能流動具有一定形狀和體積的物態)轉變為凝膠狀(不流動)的晶態。這一能引起物相變化的溫度稱為相變溫度。當環境溫度在相變溫度以上時,膜脂分子處於流動的液晶態;而在相變溫度以下時,則處於不流動的晶態。膜脂相變溫度越低,膜脂流動性就越大;反之,相變溫度越高,膜脂的流動性也就越小。
②膜脂的脂肪酸鏈:飽和程度高的脂肪酸鏈因緊密有序地排列,因而流動性小;而不飽和脂肪酸鏈由於不飽和鍵的存在,使分子間排列疏鬆而無序,相變溫度降低,從而增強了膜的流動性。脂肪酸鏈的長度對膜脂的流動性也有影響:隨著脂肪酸鏈的增長,鏈尾相互作用的機會增多,易於凝集(相變溫度增高),流動性下降。
③膽固醇:膽固醇對膜脂流動性的調節作用隨溫度的不同而改變。在相變溫度以上,它能使磷脂的脂肪酸鏈的運動性減弱,從而降低膜脂的流動性。而在相變溫度以下時,膽固醇可通過阻止磷脂脂肪酸鏈的相互作用,緩解低溫所引起的膜脂流動性劇烈下降。
除以上因素外,膜脂與膜蛋白的結合程度、環境中的離子強度、pH值等都會影響膜脂的流動性。
(2)膜蛋白的流動性
細胞膜中的蛋白質也能以側向擴散等方式運動。人們通過實驗已充分證實了膜蛋白的流動性。DavidFrye和MichaelEdidin利用螢光抗體免疫標記來測定細胞表面抗原的分布。首先用綠色螢光標記的抗體處理培養的小鼠細胞,用紅色螢光標記的另一種抗體處理人細胞。由於抗體與細胞膜上抗原的特異性結合,使得處理後的小鼠細胞和人細胞分別發出綠色、紅色螢光。在滅活的仙台病毒介導作用下,小鼠細胞與人細胞融合,所形成的雜交細胞一半呈綠色一半呈紅色。在37℃溫育40分鐘後,雙色螢光均勻分布於雜交細胞表面。螢光點的均勻分布主要是由於結合有螢光抗體的抗原在膜平面經側向擴散而重新分布。膜蛋白的運動也受很多因素影響。膜中蛋白質與脂類的相互作用、內在蛋白與外在蛋白相互作用、膜蛋白複合體的形成、膜蛋白與細胞骨架的作用等都影響和限制蛋白質的流動。
細胞膜各結構成分在膜中是不均勻分布的,結構成分的不對稱性是細胞膜的另一重要特徵,主要表現在以下方面:
(1)蛋白質在脂雙層中不對稱地鑲嵌分布。膜蛋白不同程度地嵌入脂雙層中或分布於膜表面。同時不同部位膜蛋白的種類和數量也不同。
(2)脂分子分布的不對稱性。在脂雙層中,各種類型脂分子的分布是不均勻的。一般來說,卵磷脂、鞘磷脂多分布於脂雙層的外層,而腦磷脂則多分布於內層。
(3)糖類的不對稱分布。糖類在細胞膜中的分布具有顯著不對稱性,它們只存在於膜外表面,與外層脂分子或蛋白質結合形成糖脂或糖蛋白。
