脫氧核糖核酸的複製
正文
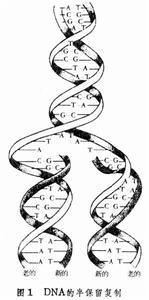
以親代 DNA為模板,合成兩個或多個與親代同樣的子代DNA分子的過程。DNA的忠實複製保證了生物性狀的代代相傳。 在體外,以DNA為模板,以脫氧腺苷三磷酸(dATP)、脫氧鳥苷三磷酸(dGTP)、脫氧胞苷三磷酸(dCTP)和脫氧胸苷三磷酸(dTTP)為底物,用DNA聚合酶可合成新生的DNA。在細胞內DNA複製是一個由多種酶協同作用的複雜過程,包括DNA複製的起動,DNA雙螺鏇的解開,新生DNA鏈的合成,複製後DNA的修飾,受損傷的DNA的修復以及複製的調節控制等方面。複製的方式 1953年J.D.沃森和F.H.C.克里克在闡明了DNA分子雙螺鏇結構之後,又提出了DNA半保留複製的假設。即親代DNA的兩條鏈可分別作為模板,各合成一條新生的DNA鏈。子代DNA中一條鏈來自親代,另一條鏈是新合成的。基於鹼基互補的原則,兩個子代DNA分子同親代的鹼基順序是完全一樣的。這個假設在1957年前後已為若干實驗所證實(圖1)。
 脫氧核糖核酸的複製
脫氧核糖核酸的複製複製叉 按半保留複製的機制,DNA分子的兩條鏈分別作為模板而進行複製,因此在複製起始點的兩端就形成叉狀結構,稱為複製叉(圖2)。
 在大多數生物中,隨著DNA複製的進行,兩端的複製叉以反向等速方式前進。某些染色體外(如噬菌體、質粒和線粒體等)的 DNA複製叉前進的方式較為特殊:或雙向而不等速前進,或僅單向前進,或以滾筒方式前進,或兩條新生鏈不同時複製。
在大多數生物中,隨著DNA複製的進行,兩端的複製叉以反向等速方式前進。某些染色體外(如噬菌體、質粒和線粒體等)的 DNA複製叉前進的方式較為特殊:或雙向而不等速前進,或僅單向前進,或以滾筒方式前進,或兩條新生鏈不同時複製。 引物 由於DNA聚合酶只能延長而不能開始新的DNA鏈合成,DNA新生鏈的合成都是以引物合成酶或RNA聚合酶先合成一小段RNA鏈為起始的,這一段RNA稱為引物。引物通常長10~50核苷酸。RNA引物最終被切除並由DNA取代。
崗崎片段 DNA聚合酶只能催化DNA鏈5'→3'方向延長,而不能催化反方向(即3'→5')的延長,因此,同複製叉移動方向一致的一條新生鏈是連續合成的,稱為前導鏈;同複製叉移動方向相反的一條鏈只能以非連續方式合成,稱為滯後鏈。崗崎首先發現這些短的非連續合成的新生片段,因此稱為崗崎片段。在引物被切除,缺口被補滿以後,崗崎片段由DNA連線酶連線成一條長鏈。
DNA聚合酶 以DNA為模板,以4種脫氧核苷三磷酸為底物,在引物存在下催化新的DNA鏈合成的酶稱為DNA聚合酶。對大腸桿菌的 DNA聚合酶研究得較為透徹。大腸桿菌包含3種DNA聚合酶,分別稱為DNA聚合酶Ⅰ,Ⅱ,Ⅲ。1956年A.科恩伯格從大腸桿菌中首次分離到 DNA聚合酶Ⅰ,因此這個酶又稱為科恩伯格酶。其特點是:①DNA模板是必需的:不存在模板時,不表現聚合酶活力;②引物也是必需的:DNA聚合酶只能從引物開始延長DNA鏈,而不能在DNA模板上重新合成一條新鏈;③以脫氧核苷三磷酸為底物,從5'→3'方向延長DNA鏈,而不催化3'→5'反應;④具有3'→5'外切酶活力:當新摻入的核苷酸同模板錯配時,它能識別並切除不配對的核苷酸,然後才能進行下一步聚合反應,因此這個酶能夠校正聚合過程中鹼基的錯配,保證DNA複製的高度精確性;⑤具有5'→3'外切酶活力:在體內DNA聚合酶Ⅰ的主要功能是切除引物,填補由此產生的缺口,並在DNA損傷修復中起作用。
遺傳學的實驗證實負責大腸桿菌染色體複製的聚合酶是DNA聚合酶Ⅲ,它具有同DNA聚合酶Ⅰ相類似的特性。在細胞內,聚合酶Ⅲ同多種肽鏈組成一個複合物,稱為DNA聚合酶Ⅲ全酶,可能它才是真正負責大腸桿菌染色體DNA複製的酶。
真核細胞中通常也含有 3種DNA聚合酶,分別稱為α、β和γ。DNA聚合酶 α存在於細胞核內,它的組成複雜而又不穩定,至今還沒有完全純化。鼠DNA聚合酶 α至少由5條肽鏈組成。同大腸桿菌DNA聚合酶不同,真核生物DNA聚合酶 α不具有3'→5'和5'→3'外切酶活力。該酶活力隨著細胞周期而變化,在DNA合成時期,這個酶活力達到高峰,因此推測這個酶負責核DNA的複製。與此相反,DNA聚合酶β的活力不隨細胞周期而變化,它的功能主要在於修復受損傷的DNA。DNA聚合酶γ可能就是早先發現的線粒體DNA聚合酶。
DNA連線酶 複製過程中必需的酶之一。 在模板存在下,它把DNA片段的5'磷酸同相鄰片段的3'羥基連線起來。在崗崎片段的連線、DNA的修復和DNA重組中,DNA連線酶都是不可缺少的。
DNA雙螺鏇的解開 一些酶和蛋白質協同作用的結果。DNA解螺鏇酶可解開複製叉之前的DNA雙螺鏇,解鏈後產生的單鏈同 DNA結合蛋白相結合而得到穩定。拓撲異構酶Ⅰ在切斷和重新連線 DNA鏈的過程中提供了一個鏇轉支點,使得由解螺鏇酶解開雙螺鏇而產生的多餘螺鏇度在這裡鏇轉而得以釋放。 拓撲異構酶Ⅱ能向DNA雙螺鏇引入負的超螺鏇,這進一步有助於雙螺鏇的解開。
DNA甲基化 DNA忠實的複製也意味著親代 DNA中甲基化鹼基的分布方式同樣複製在子代 DNA分子中。原核生物DNA中含有一些甲基化的腺嘌呤和胞嘧啶,真核生物DNA含有一定量甲基化的胞嘧啶。甲基化鹼基在DNA中的分布方式是特定的,這種特定的分布方式在真核生物中可能同細胞分化有關。根據DNA半保留複製的機制,新生的子代DNA雙鏈中的一條鏈來自親代,因此是已甲基化的,另一條鏈是新合成的,還未甲基化,所以新生DNA是半甲基化的雙鏈。甲基轉移酶以半甲基化DNA雙鏈為底物,找到甲基化鹼基的位置,並把新生鏈中相應位置的鹼基予以甲基化。也存在不需要模板的甲基轉移酶和脫甲基酶。染色體的結構可能決定它們的作用部位。
DNA損傷修復 環境中的射線,化學物質和代謝過程中產生的自由基等因素能直接損傷DNA或干擾DNA的複製過程。未修復的損傷可能導致突變,降低細胞存活能力甚至致死。DNA的損傷包括鹼基的改變或缺失,DNA鏈之間的交聯或斷裂。正常的細胞具有多種途徑用來除去損傷或不配對的核苷酸並代之以正確的核苷酸,其中主要的途徑稱為切除修復。切除修復的主要步驟是先把接近損傷部位5'端的磷酸二酯鍵切斷,切除損傷部位及其鄰近的核苷酸,利用未受損的互補鏈作為模板,合成正確的新鏈。在某些情況下,糖苷酶能識別和水解受損鹼基的糖苷鏈,然後插入酶把一個正確的鹼基轉移到空缺的糖基上去。紫外線照射可能使相鄰的胸腺嘧啶形成二聚體,有一種酶可分解這種二聚體,但需可見光,因此這個過程稱為光活化反應。如果二聚體還未被修復而複製叉已先通過了這一點,那么新生鏈上會留下一個缺口,在這種情況下,經DNA體內重組,受損傷的鏈仍可獲得正確的模板用以修復(見DNA損傷修復)。
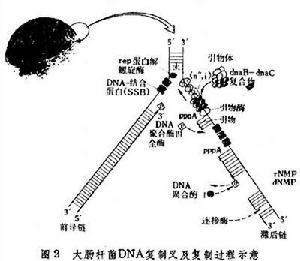
圖2總結了大腸桿菌DNA複製的基本過程。 親代DNA雙螺鏇在複製叉附近被解螺鏇酶解開,DNA結合蛋白結合在單鏈DNA上,穩定了單鏈結構,使得DNA複製能在這區域被引發並延伸。在複製叉之前產生的額外的鏇轉由於拓撲異構酶作用而釋放。結合了DNA結合蛋白的單鏈DNA同含有基因dnaB和dnaC產物的前引發複合體相結合,引物合成酶能在被引發的單鏈DNA上合成一小段RNA引物,然後 DNA聚合酶Ⅲ全酶依照模板所決定的鹼基順序從引物延伸而合成新生的DNA鏈。在滯後鏈上,每隔約1000核苷酸引發一次新生鏈。然後 RNA引物被DNA聚合酶Ⅰ的5'→3'外切酶切除,同時填滿由此而產生的缺口,相鄰的片段由DNA連線酶連線成為長的DNA鏈。以上這些酶反應順序進行,周而復始,完成整個DNA分子的複製。在其他生物中,DNA複製過程的基本規律是同大腸桿菌相仿的。
參考書目
A.Kornberg, DNA Replication,W.H.Freeman & Co.,San Francisco, 1980.
A.Kornberg, Supplement to DNA Replication,W.H.Freeman & Co.,San Francisco, 1982
