
基本介紹
李政道認為對稱性原理均根植於“不可觀測量”的理論假設上;不可觀測就意味著對稱性,任何不對稱性的發現必定意味著存在某種可觀測量。李政道說:“這些‘不可觀測量’中,有一些只是由於我們目前測量能力的限制。當我們的實驗技術得到改進時,我們的觀測範圍自然要擴大。因而,完全有可能到某種時候,我們能夠探測到某個假設的‘不可觀測量’,而這正是對稱破壞的根源。
這和“對稱性破缺則是由‘巨觀’走向‘微觀’而展現事物差異性的方式”哲學觀點是一致的。
假如沒有對稱性破缺,這個世界將會失去活力,也將是單調、黯淡的,也不會有生物。自然界同樣也存在著諸多對性破缺的例子。比如:弱作用力下的宇稱不守恆、粒子與反粒子的不對稱、手性分子的對稱性破缺等等。
系統
耗散理論在解釋生命分子手性起源中取得了較大成功,這也是本書所擁護的觀點;近些年也得到更多的實驗支持。普利高津(Prigogine)認為,在遠離平衡的條件下,一個開放的物理化學體系可以通過分支現象,從原先空間均勻的各向同性狀態發展到集中都是穩定的但時空特性可能不同的有序狀態,即由無序中產生有序。這兩種空間有序狀態唯一的差別可能僅僅在於其對稱性,體系遠離平衡態時在分支點附近對微小擾動是敏感的。1998年Kenso Soai和他的小組證實了分叉結構(bifurcation framework)。他們採用了混合了對映異構的亮氨酸,其中一種構型少量過剩。在這不均衡的溶液中反應形成的嘧啶醇也有一種對映體少量過剩。這種分子在自身形成過程中能起催化作用,因而占主導地位。因為自催化的循環反應結構放大了這個細微手性破缺效應,這使得生命分子為擇單一手性。
1995 年3 月,美國《科學》雜誌報導在洛杉磯召開的“生物分子手性均一起源”的國際會議上,與會的物理、化學、天文學家大多數認為,“沒有手性就沒有生命”,“手性起源先於生命”而不是生命自然選擇了手性。
2006 年6 月1 日出版的《Nature》7093 期第621 頁一篇題為“Thermodynamic control of asymmetric amplification in amino acid catalysis”的文章,Martin Klussmann 等人提出了手性分子不對稱擴增的另一種解釋,這是對自催化機制的一種替代機制。與傳統手性藥物合成不同的是,這是一種動力學控制下的不對稱擴增。
總的來說分子的手性根源來自於弱相互作用,這說明電磁作用力並不是化學尺度上唯一影響化學分子的作用力。弱相互作用所誘發的費米子的手征性,主要通過反饋特徵的自催化效應及其它某种放大機制,放大到分子尺度,從而成為生命形成重要的驅動因素。
物理
物理學中幾何對稱與抽象對稱
對稱性破缺可以理解為原來具有較高對稱性的系統,出現不對稱因素,其對稱程度自發降低的現象。或者用物理語言敘述為:控制參量λ跨越某臨界值時,系統原有對稱性較高的狀態失穩,新出現若干個等價的、對稱性較低的穩定狀態,系統將向其中之一過渡。和前面群論提到幾何對稱操作中旋轉、反映、反演相似,在物理學中則是電荷對稱、時間反演、空間反映,的對稱操作就是C、T、P。CTP也存在對稱與破缺。
按照諾特定理,守恆量意味著對稱性;在物理學上不僅僅有幾何的對稱還有抽象的對稱。比如:電荷守恆定律涉及抽象的性質而非動力學的性質,它對應著抽象的對稱性;還有保守力在保守場中的做功,這些就是規範對稱。在尋求各種相互作用力的理想的量子理論中,規範對稱性在起著核心的作用;而且統一力的理論嘗試也是在規範對稱性的範圍之內的。
 對稱性破缺 對稱性破缺 |
超對稱
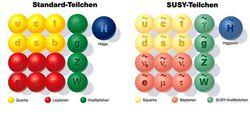
在70 年代早期,理論物理學家發現比旋轉和平移這種操作更深刻,更有效的幾何對稱性,這就是超對稱。比如具有1/2 費米子必須轉動720°才能回復到原先的位置,這種雙值性決定了它的幾何對稱操作玻色子完全不同。這樣的性質在通常空間的幾何操作的框架內是不能處理的。在超對稱理論中,在通常的四維時空上附加另外的四維,稱為超空間,目的是為了容納費米子奇異的幾何性質。因此這些附加的“費米維數”不是我們所知的空間或時間的維數。超對稱操作能從通常空間轉到附加的費米維數,即能把玻色子變為費米子,把費米子變為玻色子。所以我們可以把費米子和玻色子看成一個幾何存在的兩個不同的投影。
 對稱性破缺 對稱性破缺 |
按照這個理論,費米子和玻色子具有直接的物理聯繫,費米子與玻色子相互對應,即每個粒子都有其超對稱伴侶;但從現在已知的玻色子和費米子來看,這種對稱性似乎不成立的。有觀點認為,現在物理學中系統的對稱性破缺,是因為更深層次的對稱性是隱藏著的,也許自然本身是超對稱的;同樣,超對稱也會出現真空自發破缺。
德國數學家卡魯扎(Kaluza)提出,通過附加一維額外的虛空間自由度來寫出五維而不是四維的愛因斯坦引力場方程;五維的愛因斯坦場方程不但給出了通常的四維引力方程,還給出另外一組方程,而這正好就是電磁場的麥克斯韋方程組。按照這個理論電磁作用和引力作用都不是單獨的力,而是在不可見的更高維空間自由度的世界裡。奧斯卡·克萊因(O.Klein)認為我們沒能察覺到那一維額外自由度的原因是,在某種意義上它“捲縮”到一個非常小的尺度,就好比一個水管在遠處看就會把它當成一個曲線。這個尺度就是普朗克尺度,10*(-35) 米是空間被分割的最小距離。
弱作用規範
史蒂芬·溫伯格(Steven Weinberg)和阿卜杜斯·薩拉姆(Abdus Salam)各自獨立地發現有可能在不破壞弱作用內在的規範對稱性的情況下使弱“媒介”粒子獲得質量。這一質量可以通過弱作用場內部一定的相互作用來自發地產生,弱作用的規範對稱性可能是自發破壞而不是動力學破壞。整體對稱性是一個連續變換群,整體對稱性自發破缺,零自旋、零質量粒子就會產生,稱為戈德斯通(Goldstone)玻色子,如果局部對稱規範群自發破缺,部分戈德斯通玻色子將會得到質量,即希格斯機制。溫伯格和薩拉姆提出W 和Z 粒子(弱作用的“媒介”粒子)是通過弱作用希格斯機制獲得質量的。希格斯場量子是有質量無自旋的玻色子,它與電磁-弱作用場相耦合,在這種耦合的作用下,系統選擇了最低能量狀態,使得W 和Z 獲得大質量。
根據超對稱理論,暗物質粒子稱為neutralinos ( 常稱為WIMPS),彼此湮滅釋放次級粒子和輻射,包括中等能量伽瑪射線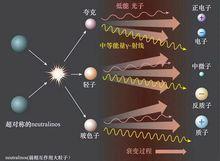 對稱性破缺 對稱性破缺 |
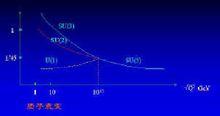
對規範對稱的描述也要用到數學的群論,描述這種連續對稱的稱為李群。例如圓環上的對稱性,一個圓環在繞其中心軸轉動任何角度時保持對稱。這些轉動構成一個群,稱為U(1),其中U 代表“么正”的意思,是一種特定的數學性質。碰巧電磁場的規範對稱性正是這種U(1)對稱,為Able 群;不過是在某一抽象空間中,而非真實的空間。弱力和電磁力可由SU(2)xU(1)非阿貝爾規範理論來統一描述,S 代表“特殊”;已有標準模型:SU(3)x SU(2) x U(1)(非阿貝爾規範理論)來描述強、弱、電磁三種力,我們在這裡並不關心它在數學上的具體含義。
 對稱性破缺 對稱性破缺 |
11維空間
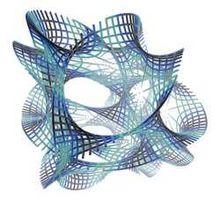 M-理論中11維空間示意圖
M-理論中11維空間示意圖上面提到超對稱幾何也可以作為引力幾何理論的基礎,相應的理論就稱為超引力論。在超引力中,引力子已不再是傳遞引力的唯一媒介粒子,超對稱是在費米子與玻色子之間提供了某種聯繫。按照這個理論應該存在一種自旋3/2 的基本粒子,稱為引力量子。引力量子的超對偶粒子總數達172 個,由於這一理論中有8 種引力量子,人們稱之為N=8 的超引力理論。劍橋大學的史蒂芬·霍金(Stephen Hawking)在他就職Lucasian 數學講座教授的演說中提到,假使N=8 的超引力大有希望,那么“理論物理的終結為期不遠了。” 當理論在大於四維的時空中構造時,超引力的幾何結構可以大大簡化,對於N=8 的超引力,最為有利的維數是11。在11 維的克萊因-卡魯扎理論中,只存在一種力--引力,而電磁,弱和強力只不過是引力的附屬品。也許我們就在11 維空間中,所以筆者叫做網名11 維空間,便由此得來。
11 維空間能否成立? 弱作用的一個明顯特徵是它破壞左右鏡象對稱,這意味著基本粒子具有‘手性’,確定的手征性只存在於單數維的空間中;也就是說,空間的維數是奇數,因而時空的維數必定是偶數。這是11 維空間遇到的難題。
生物
對稱性破缺與生物起源
二次疊代Mandelbrot 集與分子填充為組織的疊代比較。他們源於不同的非線性過程,諸多分形結構都具有M 集的類似特徵。這個過程是高度非線性的,依賴這樣的非線性使得結構具有自相似性。生物源於宇宙對稱性破缺,分子系統豐富多彩的結構也通過對稱破缺而發展。
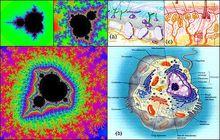 對稱性破缺 對稱性破缺 |
那么從宇宙物質產生到產生生命要經歷那些不對稱的過程呢?
宇宙形成過程就是對稱性破缺,最初的宇宙為對稱真空態(作為奇點的量子真空以指數方式膨脹,即暴漲),宇宙的對稱性逐步喪失。隨著宇宙的膨脹和降溫,原真空態發生一系列相變:10-44s 時,引力作用分化出來,夸克和輕子可相互轉變。
 對稱性破缺 對稱性破缺 |
10*(-36)s 時強相互作用同電,弱作用分離,開始出現物質與反物質的不對稱;10*(-10)s 後,弱作用與電磁作用分離。要產生物質構成的世界,就必須正反物質不對稱;否則就會湮滅。最初宇宙正反物質幾乎一樣多,不對稱度僅為A=3×10*(-8)。宇宙演化產生不對稱機制,使得重子數不對稱;這就要求CP的對稱破缺,打破了重子數的守恆,從而形成現在物質占主要的世界。物質在四種作用力下分層次演化。通過複雜的核合成過程形成早期的原子核,再形成原子和分子。
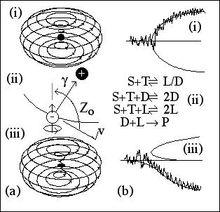 手性弱力提供了對稱破缺的擾動
手性弱力提供了對稱破缺的擾動前面提到電弱作用力間對稱破缺的Higgs 機制,在最低能量下不為0 的真空極化從而導致電弱對稱破缺。電弱作用力通過“弱荷流”(W)和“弱中性流”(Z )區分的(弱荷看成電荷,中間玻色子看成電磁波的光子)。任何兩個基本粒子間的這些流的強度取決於粒子間的距離和它們的電荷。電子檔靠近原子核時,受到中性弱力引起電子軌道的擾動,使它選擇手性。如圖(i)軌道是非手性的,但是由於Z0 擾動導致出現手性旋轉,圖中(b)代表自發對稱破缺(Autocatalytic symmetry-breaking)源於隨機的手性分岔,弱擾動破壞穩定性從而形成具有手性的(iii)。這樣的對稱破缺會被放大到聚合系統中,在分子D 和L 構型的競爭中,哪個具有負反饋系統那個就會占主導。
奧克蘭大學的Chris C. King 在宇宙生物學論文中提到在Murchison 隕石(1969 年落於澳大利亞)上非生物胺基酸往往傾向於左手構型。這說明,在生物產生以前這種不對稱機制就可能已經存在。
手性破缺
宇稱不守恆弱相互作用對應於產生手性分子有兩種假說。一種是以帶電電流宇稱不守恆(Charged Current-Parity Non conservation:CC-PNC)為基礎的假說,認為由於CC-PNC,β衰變產生徑向極化電子,進入物質內部產生圓偏振光輻射,導致底物分子立體選擇合成或分解,從而產生手性分子。
另一種以弱中性流宇稱不守恆(Neutral Current-Parity Non conservation:NC-PNC)為基礎的假說,認為NC-PNC 造成對映體之間微小的能差叫宇稱破缺能差(Parity Violating Energy Difference, PVED),PVED 進一步在對映體分子的物理性質及反應速率上形成差別,經過放大可以產生均一的手性。
Vester-Ulbricht機理一個基本粒子(電子或正電子)靜止時是球對稱的,因此是非手征性的。但一個自旋粒子沿著自旋軸的任一方向移動時,它就成為手征性的。β-電子為左手螺旋電子,β+電子為右手螺旋電子。Goldharber 在1957 年發現,從某一輻射核ß 衰變中產生的電子是徑向偏振的,由此電子產生的韌致輻射產生圓偏振光。
1959年Vester 和Ulbricht首先將β衰變的不對稱性和生物分子的不對稱性聯繫起來,提出Vester-Ulbricht機制。不對稱性可從基本粒子水平轉移到分子水平,ß 衰變所表現的手性與自然界生物分子的手性間有因果關係。ß 衰變中產生的偏振電子,對外消旋分子或前手性分子產生非對稱性影響,最後由不對稱合成或分解產生手性分子。β衰變→縱向偏振電子→園偏振電子→不對稱光化學作用→手性的。
Vester-Ulbricht機理的相關實驗1968 年,Garay 將1.33e7Bq 的SrCl2,分別加入到L 與D 型酪氨酸的鹼性溶液中,在室溫放置18 個月後,發現D-酪氨酸分解比L 型顯著,並認為這是由於發生偏振光的ß 射線作用,是D-酪氨酸進行了立體有擇性分解。1975 年Bonner 用120keV 的線性加速器,來產生天然的自然反平行(AP)“左手性”電子,也可產生非天然的自旋平行(P)“右手性”電子,進行輻解D-和L-亮氨酸的實驗。在53%-76%樣品分解後,AP 電子產生0.60%-1.42%過量的L-亮氨酸,P 電子可產生0.74%-1.14%過量的D-胺基酸。1976年Darge 等將標記32P 的磷酸鹽加入到消旋的D、L-色氨酸水溶液中,在-25℃下放置12 周后測定紫外光譜,發現色氨酸的分解率為33%,必旋光度為+(0.7±0.4)e-3 度。由此可見L-色氨酸比D-色氨酸優先分解,造成了大約19%的D-色氨酸的濃集。這一結果與Garay的結果矛盾。但Bonner重複Garay和Darge的實驗,Hodge重複Bonner的實驗,都沒得到立體選擇的結果,使得這一假說爭論了近半個世紀。
王文清認為γ射線無手性選擇分解;β電子對胺基酸的不對稱分解只有在低溫、初始階段遠離平衡態,即在開放體系才能得到,如無放大機制,達到熱力學平衡差別消失。王文清根據王建英、羅遼復理論研究,以量子力學的理論方法,從β電子與手性分子的電磁作用出發,探討了β電子和正電子在不對稱分子上的非彈性碰撞,證明了對於對映異構體D-和L-胺基酸分子,碰撞截面的相對差值。差值F與旋光強度和和偶極強度的比值成比例,數值上為10-6量級,通過運算得到,當L型分子旋光強度大於零時,極化電子優先分解D型分子,反之當旋光強度小於零時,極化電子優先分解L型分子。1993 年王文清、羅遼復在義大利國際生命起源會議上指出:長期以來,人們忽視了構型和旋光興並不是同一概念,ß 粒子是對胺基酸的旋光性(左旋Rn<0,,右旋Rn>0)有選擇,而不是對構型(D,L)有選擇。
β電子和手性分子左右不對稱碰撞,導致某些特定的化學反應速率常數的不對稱為10*(-6)量級,並且對於每一確定的化學反應都有確定的符號。速率常數的不對稱性將在反應擴散方程中加入不對稱的外力項,在反應擴散方程的解-L和D型分子的濃度中,這種不對稱效應被明顯放大。根據丁達夫、徐京華分析,如果不對稱外力為η量級,則解的不對稱性可達η1/3的量級。因此,在β電子照射下,通過適當的化學反應可使D和L型胺基酸分子的相對濃度差別達到10*(-2)~10*(-3)。另一方面,如果這種化學反應與多聚鏈的形成有關,當鏈長為10*(2)~10*(3)個分子時,L和D型分子鏈的濃度差別將達到0(1)的量級。這就有可能解釋為什麼很多生物大分子都具有確定的手性。
Salam 假說
1991 年Salam 提出:Salam認為電磁力不是唯一引起化學反應力,電弱Z0也在化學效應中期作用。由於Z0相互作用,電子與電子耦合成庫柏對,藉助量子力學協同效應,由於玻色凝聚,在某一臨界低溫Tc 下引起二級相變,包括D 型胺基酸向L性相變。一般來說,Tc 是個低溫值,地球作為L-胺基酸形成之地太熱了。所以他構想,在低溫、原始宇宙空間早在地球形成前胺基酸的手性選擇就已進行(這和隕石發現吻合)。S.F.Mason G.E.Trantar 對若干個L 型和D 型胺基酸的能量進行了詳細的計算,並考慮了不對稱的Z力,預料中的對映體之間的能量分裂出現了。在所有情形下,生物學上占優勢的L-胺基酸和D-核糖都具有較低的能量。計算結果如下:(L-D)Ala:
-3.0×10-19ev;(L-D)Val:-6.2×10-19ev;(L-D)Ser:-2.3×10-19ev;(L-D)Asp:-4.8×10-19ev;(L-D)核糖:+1.8×10-19ev可見,對映體分子間的能量差數量級為10-19ev。300K 時,L型比D 性胺基酸數量多1/107。
1994 年4 月王文清、盛湘蓉與楊宏順、陳兆甲科研組合作,利用搽粉絕熱連續加熱量法在77.35K 到300K 區間,以0.5K/min升溫速率(或降溫速率)測定D-纈氨酸和L-纈氨酸的比熱容與T 圖,發現D-纈氨酸在270±1K 有明顯λ相變,而L-纈氨酸則無。經多次熱循環及以L-纈氨酸作參比樣品,D-纈氨酸在同一溫度均重複出現比熱容尖峰。D-纈氨酸單晶X 衍射晶格數據顯示,在臨界溫度Tc 前後,無明顯晶格變化。試探排除了水汽、結晶水及晶格變化對比熱容的貢獻,王文清等人認為比熱容異常是由於電子耦合成庫柏對及協同效應的S電子比熱容貢獻,並認為D-纈氨酸的相變可能是D 型向L 型轉變的二級相變。同樣,對於D-丙氨酸單晶分子,在低溫200K 到300K 出現了磁相變,這與比熱容測出的λ相變溫度一致,人們認為,該溫度也許就是D-丙氨酸向L-型轉變的二級相變。
Salam 假說:1.亞原子水平上表現出的Z 力結合電磁相互作用,在凝聚態胺基酸單晶D 和L 型分子中被首次檢測到,這在理論上有重大意義;2.D-丙氨酸和D-纈氨酸均在實驗中發現了比熱容的λ相變,這將有助於證明Salam 假說中提到的D 型胺基酸向L-型轉變的二級相變。
局限性
首先Garay等人的實驗都不能嚴密地證明V-U機理,Bonner 的實驗中,當電子能量為60eV 時,選擇分解消失。而且,對上述實驗的一些重複也往往既得不到肯定也得不到否定的結果。按照王文清提出的構型和旋光興並不是同一概念,ß 粒子是對胺基酸的旋光性有選擇,而不是對構型(D,L)有選擇。因此不能解釋生命分子構型的手性單一性。而在Salam 假說中10-19ev如此微小的機率差異,能否成為生命選擇單一手性機制的原因?就算二級相變存在,從胺基酸混消旋體轉變成某種手性單一的胺基酸分子的過程,需要越過一個大的活化能勢壘。如果這個相變發生,將導致化學鍵斷裂。顯然,這兩種學說及相關實驗只證明了手性分子極其微小的對稱差異,這不是生命手性分子起源的根本。隕石分子的發現和這些實驗說明在生命產生之前,手性分子在數量上微小的差異就已經存在;但這樣的差異必定要通過一個放大機制給予放大,才能形成現在生物分子的單一手性。
耗散分岔
耗散理論在解釋生命分子手性起源中取得了較大成功,這也是筆者所擁護的觀點;近些年也得到更多的實驗支持。普利高津(Prigogine)認為,在遠離平衡的條件下,一個開放的物理化學體系可以通過分支現象,從原先空間均勻的各向同性狀態發展到集中都是穩定的但時空特性可能不同的有序狀態,即由無序中產生有序。這兩種空間有序狀態唯一的差別可能僅僅在於其對稱性,體系遠離平衡態時在分支點附近對微小擾動是敏感的。1998年Kenso Soai和他的小組證實了分叉結構(bifurcation framework)。他們採用了混合了對映異構的亮氨酸,其中一種構型少量過剩。在這不均衡的溶液中反應形成的嘧啶醇也有一種對映體少量過剩。這種分子在自身形成過程中能起催化作用,因而占主導地位。因為自催化的循環反應結構放大了這個細微手性破缺效應,這使得生命分子為擇單一手性。
1995 年3 月,美國《科學》雜誌報導在洛杉磯召開的“生物分子手性均一起源”的國際會議上,與會的物理、化學、天文學家大多數認為,“沒有手性就沒有生命”,“手性起源先於生命”而不是生命自然選擇了手性。
2006 年6 月1 日出版的《Nature》7093 期第621 頁一篇題為“Thermodynamic control of asymmetric amplification in amino acid catalysis”的文章,Martin Klussmann 等人提出了手性分子不對稱擴增的另一種解釋,這是對自催化機制的一種替代機制。與傳統手性藥物合成不同的是,這是一種動力學控制下的不對稱擴增。
總的來說分子的手性根源來自於弱相互作用,這說明電磁作用力並不是化學尺度上唯一影響化學分子的作用力。弱相互作用所誘發的費米子的手征性,主要通過反饋特徵的自催化效應及其它某种放大機制,放大到分子尺度,從而成為生命形成重要的驅動因素。
反饋機制
我們知道 生命分子的產生是源於反饋的自催化機制通過循環結構將微小的差距放大;而我們的社會也是一個充滿張力的循環結構。自然界存在各式各樣的不對稱差異,能夠放大這樣差異的則是事物自身反饋效應。在達爾文進化論中性別選擇是一個很重要的部分,這可以理解為物種自身對自身的一個選擇力,也具有反饋特徵。
高等動物進化出來的互相扶持以及護幼行為等都是基於群體意識,這也是物種對自身的反饋,簡單的說就是“自我選擇力”。中國儒家傳統思想所尊崇的信條就是以自我完善為基礎,在《禮記·大學》中就有“心正而後身修,身修而後家齊,家齊而後國治,國治而後天下平。”這樣的思想是符合生物哲學的,人的修身必須從自我反饋開始。然而現今中國的教育,卻沒有教會人,適應和反饋這最重要的東西。引用卡內基梅隆大學教授藍迪的“最後一課”的演講中的一句話“一個教育工作者能給的最好禮物,就是讓人能自我反省”。
舉例
下面列舉幾個對稱性自發破缺的事例:
宇稱不守恆
 對稱性破缺
對稱性破缺實驗已經證明,強作用下宇稱守恆。這是與微觀粒子的鏡象對稱性相聯繫的守恆定律。1956年前後,在對最輕的奇異粒子衰變過程的研究中遇到了“t ~ q 疑難”。實驗中發現的t 和q 粒子,它們質量相等,電荷相同,壽命也一樣。但它們衰變的產物卻不相同:實驗結果的分析表明,3個p 介子的總角動量為零,宇稱為負。而2個p 介子的總角動量如為零,則宇稱只能是正。因此,從質量、壽命和電荷來看, q 和t 似乎是同一種粒子。但從衰變行為來看,如果宇稱是守恆量,則q 和t 就不可能是同一種粒子。
1956年,李政道和楊振寧解決了這個難題。他們提出弱相互作用過程中宇稱不守恆的構想,吳健雄的鈷60原子核b 蛻變實驗驗證了這個構想。1957年,吳健雄在10-2 K下做原子核b 衰變實驗,用核磁共振技術使核自旋按確定方向排列,觀察b 衰變後的電子數分布,發現無鏡像對稱性 —— 證明了弱作用的宇稱不守恆性。
1957年李政道和楊振寧獲諾貝爾物理獎。
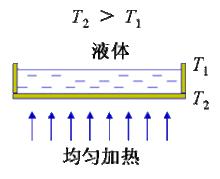 對稱性破缺
對稱性破缺貝納德對流
1900年法國學者貝納爾 (H.Benard)發現:從下面均勻加熱水平容器中薄層液體時,若上下溫差超過一臨界值, 液體中突現類似蜂房的六邊形格線, 液體的傳熱方式由熱傳導過渡到了對流,每個六角形中心的液體向上流動,邊界處液體向下流動。這是對流與抑止因素(黏性和熱擴散)競爭的結果。
 對稱性破缺
對稱性破缺義大利怪鍾
這是1443年 Paolo Uccello繪製的24小時逆時針方向運行的“怪鍾”(如圖)。經濟學家Arthur Brian以此鍾為例,論述經濟領域中的正反饋現象。他說,1443年鐘的設計尚未定型。一種錶盤的設計用得愈多,就有更多人習慣於讀它,以後它就被採用得愈多。最後形成現在的慣例。這就是從 正反饋到失穩,再從失穩到對稱破缺的過程。
重子與反重子
1933年Dirac理論預言: 每種粒子都有自己的反粒子, 正反粒子完全對稱,也許在遙遠的地方存在“反物質世界(anti-world)”。按照粒子物理學的分類,質子、中子以及它們的反粒子都屬於重子,重子數B 是個守恆量。重子數 B 的定義是:每個重子的B =1, 每個反重子的B =-1。於是,在重子對產生和湮滅的過程中,重子數總和保持為零。各種天文觀測表明: 宇宙線中反質子與質子數量之比< ;無論在太陽系內、銀河系內、還是整個星系團的更大範圍內,都未觀察到湮沒引起的強大g 射線。如果認為重子數守恆是一條在任何情況下都顛撲不破的定理,就只好認為,宇宙從它誕生時刻起就存在現今那樣多的不為零的重子數,即重子與反重子一開始就不對稱。目前,對正、反重子不對稱比較可能的解釋是,早期極高溫的宇宙中存在著違反重子數守恆的過程。
生物界套用
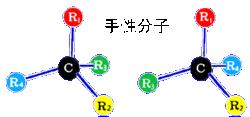 對稱性破缺
對稱性破缺大多數動物在外觀上都具有左右對稱性,但體內的器官就不那么對稱了。如果深入到分子層次,就會發現一種普遍存在於生物界的更深刻的左右不對稱性。1844年德國化學家E.E.Mitscherlich發現,酒石酸鈉銨和葡萄酸鈉銨的結晶具有相同的晶形,一樣的化學性質,但溶液的旋光性不同。前者使偏振面右旋,後者無旋光性。1847年法國Louis Pasteur發現了葡萄酸鈉銨中有互為鏡象對稱的兩種旋光異構物,其結構如圖所示。對此現象解釋的信念是:光活性有與生命過程相聯繫的起源。
現代生物化學指出:有機化合物的旋光異構現象與有機分子中碳原子四個鍵的空間構形有關。用L(livo)和D(dextro)分別表示左、右型旋光異構體,(+)、(-)代表該物質的溶液的旋光方向,(-)表示左旋,(+)代表右旋。碳四面體的左右兩種構型、甘油醛中四個基團L、D兩種構型以及丙氨酸的旋光異構體簡要圖示如左圖,它明顯地反映出了其結構的左右不對稱性。生命的基本物質是生物大分子,它包括蛋白質、核酸、多糖和脂類。其中蛋白質是生命功能的執行者,其分子是右胺基酸組成的長鏈。每種胺基酸都應有L、D兩種旋光異構體。但實驗證明組成生物蛋白質的20種胺基酸都是L型的,D型胺基酸只存在於細菌細胞壁和其它細菌產物中。核酸是遺傳信息的攜帶者和傳遞者,分為核糖核酸(RNA)和脫氧核酸(DNA)兩種。右下圖是DNA分子雙螺旋結構模型,通常是右旋的。這正是生物大分子的手性特徵。生物體內化合物的這種左右不對稱性正是生命力的體現。維持這種左右不平衡狀態的是生物體內的酶,生物一旦死亡,酶便失去活力,造成左右不平衡的生物化學反應也就停止了。由此可見,生命與分子的不對稱性息息相關。問題是地球上生命發源之初,左右對稱性的破缺是怎樣開始的?即分子手性的起源是什麼?生物的起源是什麼?這些都是有待人們去研究的謎。
總之,時空、不同種類的粒子、不同種類的相互作用、整個複雜紛紜的自然界,包括人類自身,都是對稱性自發破缺的產物。對稱性破缺的機制是什麼?實在現象中的對稱性破缺與基本物理規律的對稱性是否相容?不同層次的非對稱性間如何關聯?這些都是現代物理尚未解決的重要課題。
真空不空
宇宙廣大區域的真空中運行著光速的光子、中微子,超光速的引力子、反引力子,用E1=ma2方程計算,真空中蘊藏著的能量是很大的,而且不同區域的真空蘊藏的能量差異極大,如黑洞奇點的真空區和宇宙奇點的真空區與宇宙廣大區域的真空相比較。
宇宙真空充滿了引力子和反引力子,而且由於純引力的黑洞存在,宇宙總體上已出現了引力子和反引力子的不對稱,即引力子總量多於反引力子。對稱性破缺的本質來自於宇宙真空的不對稱性產生真空對稱性自發破缺機制。
如果系統受到一個小擾動破壞了它的對稱性,我們說它的對稱性破缺,比如,原子中的這樣一個擾動可以由電場引起,由於擾動的作用,原子將不再停留在它原先的定態上,而從一個能級躍遷到另一個能級,並發射或吸收一個可見光光子。對稱性破缺同樣出現在粒子中,這時的干擾因素就是宇宙中無所不在的引力子和反引力子。之所以出現“宇稱不守恆”,是因有些粒子在真空中的引力子、反引力子的干擾下,必然會出現上述現象,而且較易出現在有弱核力參與的粒子轉化過程中,因為這種力較弱,即反引力場較弱,較易受到外界的引力子或反引力子的干擾。
對稱性破缺也叫CP破缺
 對稱性破缺
對稱性破缺在宇宙中,上下級物質特別容易產生干擾,形成對稱性破缺,粒子級物質較易對原子形成干擾,因為前者是後者的結構材料,同理,引力子級物質較易對粒子形成干擾,形成對稱性破缺。而引力子級物質對原子、分子、生物體較難在短期內形成可察覺的干擾,因為它們存在巨大的質量差異,這種干擾只能漸進式的,一種從“量變到質變”的緩慢過程,引力子級物質最先影響粒子級物質,通過它逐漸對原子形成影響。
粒子世界的“不確定”、“測不準”就是因為粒子質量太小,而宇宙真空中的引力子、反引力子密度比光子、中微子等粒子高出很多倍,引力場使得巨觀宇宙的時空都發生彎曲,粒子在無數引力子和反引力子的碰撞干擾下,出現“不確定”、“測不準”是必然的。
正是真空的這種特性,造成“宇稱不守恆、CP破壞及時間(T)反演不變性的破壞、規範對稱性的自發破缺”等一系列對稱性丟失。而且宇宙必須存在對稱中的不對稱,完全對稱的宇宙將會凝結,如果正奇子與反奇子在對抗與協同中完全對稱,將不可能形成引力子與反引力子,如果正、反夸克組合出完全對稱的正、反質子,正、反中子,今日的宇宙將只剩下微波輻射。
社會
反饋機制與社會
生命分子的產生是源於反饋的自催化機制通過循環結構將微小的差距放大,社會也是一個充滿張力的循環結構。自然界存在各式各樣的不對稱差異,能夠放大這樣差異的則是事物自身選擇。同樣在達爾文進化論中性別選擇是一個很重要的部分,這可以理解為物種自身對自身的一個選擇力,也具有反饋特徵。
高等動物進化出來的互相扶持以及護幼行為等都是基於群體意識,這也是物種對自身的反饋,簡單的說就是“自我選擇力”。中國儒家傳統思想所尊崇的信條就是以自我完善為基礎,在《禮記·大學》中就有“心正而後身修,身修而後家齊,家齊而後國治,國治而後天下平。”這樣的思想是符合生物哲學的,人的修身必須從自我反饋開始。這讓我想起,美國電視《越獄》中有一句話“欲變世界,先變其身”。然而現今中國的教育,卻沒有教會人,適應和反饋這最重要的東西。引用卡內基梅隆大學教授藍迪的“最後一課”的演講中的一句話“一個教育工作者能給的最好禮物,就是讓人能自我反省”。
生活中學會總結,是人生自我反饋的開始。社會上每個人都是不同的,自然屬性賦予了人差異性的一面,只有自身對自身的反饋來放大這種差異,人生才會精彩(這包括自我修養和自我超越)。自古封建君王們都鼓吹‘君權神授’;也是企圖放大,人的的差異,將自我比作神。而現代社會人在置身於物慾世界的同時,忽略了自我對自我的反饋,盛行的卻是類似斯賓塞弱肉強食的“社會達爾文主義”。
社會達爾文主義忽略了社會中事物發展自身反饋也是重要重要驅動,具有局限性,因此被後現代主義稱為“現代性罪狀”。在這樣的扭曲的社會結構中,人們追求自我實現,不是通過自我修養和超越的反饋來完成;而以掌握物質財富和社會地位來衡量,力求成為社會“食物鏈”的頂端。同時,在張力的社會中人們文化的困境與內心的掙扎也是推動其發展的驅動因素。在霍妮的文化心理病理學指出自我的掙扎是人與自我關係的失調。人有天賦的潛能和引導實現潛能的建設性力量,體現為創造和奉獻;這種力量的激發則需要人自身的“自催化”,其過程是通過學習、經歷、以及自我認識來完成。
同時人的天賦中還具有一種破壞性力量,體現為貪婪、權利與欲望的膨脹等等。為確保社會結構穩定,需要社會機制的約束和自我反饋加以調節,這表現為法律與道德。一些人認為這種破壞的力量歸結為人類的本能,其實這是片面的,人類的本性是兩種力量的綜合,而不是單純某一方面。就以‘性’來說,弗洛伊德的人性論是性惡論,並持悲觀論調;但我們知道‘性’又意味著生命的誕生,意味著創造,意味著美,具有積極的一面。
人能夠調節這兩種力量的就是自我的反饋,並體現為適應性。生物要適應環境得以生存,就首先要求自身的改變,這個變化過程就是自身反饋機制的體現。反社會人格以及神經症患者內心的掙扎以及自我異化等,在我看來是社會適應力低下的表現,可能是自身反饋出現了問題;按照這個思想,極度自卑或自傲都可能滋生反社會行為;我相信運用這個思想是可以找到減少社會暴力的方法。當然,社會是多元化整體,事物的發展既取決於自身反饋又取決於環境的選擇。假如社會環境變化總採取突變式,或者說環境選擇的跳躍變化總大於自我反饋的能力,那么這樣的反饋機制就可能遭到破壞。所以在社會學中人自我的反饋機制往往具有強烈的環境依賴性。
假如構成社會的人,都具有極強的適應能力,都在不斷的變通;那么這個社會是不穩定的,比如可能社會缺乏誠信、缺乏價值判斷等等。所以社會本身是人社會適應性與社會穩定性的妥協。而在生態學中生物與環境本身就是一個整體,是協同進化的。一個物種的進化,既有來自外界的自然選擇驅動;又有物種內的性別選擇自身反饋。環境的變化必定帶來物種自身反饋的變化,所有物種自身的變化又會積累從而改變環境。環境是超循環結構,其穩定性與物種的多樣性密切相關,因此多樣性的環境是有能力抵禦少數物種突變對環境的改變。生物圈本身就是一個物種適應性與環境穩定性的妥協;而生物個體就是自我適應於自我結構穩定的妥協。
