
特徵
 埃德蒙頓龍化石
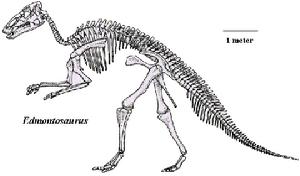
埃德蒙頓龍化石完全成長的埃德蒙頓龍可達9米長,一些較大的標本則達12米[5]到13米長。體重約4.0噸,是最重的鴨嘴龍科之一。帝王埃德蒙頓龍是體型最大的種;在2004年,傑克·霍納(Jack Horner)等人提出科氏大鴨龍是E. annectens的異名,但很少其他研究引用這個分類法。模式標本(編號NMC 2288)的身長估計為9到12米。E. annectens已發現兩個標本,身長分別為8米、8.92米。薩斯喀徹溫埃德蒙頓龍的身長則為7到7.3米。
埃德蒙頓龍的頭骨長度約1米,帝王埃德蒙頓龍的頭骨較長,而E. annectens的頭骨較短。埃德蒙頓龍的頭部側面略呈三角形,而且沒有骨質頭冠。若從頭骨的上方看,頭部的前端與後段較寬,中段狹窄。埃德蒙頓龍具有缺乏牙齒的喙狀嘴,由角質組織覆蓋。根據位在森肯貝格博物館的“糙齒龍木乃伊”標本,埃德蒙頓龍的喙狀嘴角質部分至少有8厘米長。埃德蒙頓龍的鼻孔大,鼻孔周圍的骨頭凹陷。根據“糙齒龍木乃伊”標本,埃德蒙頓龍的眼眶具有鞏膜環(Sclerotic rings)。
 埃德蒙頓龍
埃德蒙頓龍埃德蒙頓龍只有上顎骨與齒骨具有牙齒。新的牙齒會不斷的生長來取代脫落的牙齒,一顆牙齒需要約一年的時間來生長。牙齒排列成形成數十列齒系(Dental batteries),每列齒系有至少6顆牙齒;而齒系的數量,依該物種而變動。帝王埃德蒙頓龍的每塊上顎骨有51到53列齒系,每塊齒骨有48到49列齒系,而上顎的牙齒較下顎牙齒狹窄。E. annectens的每塊上顎骨有43列齒系,每塊齒骨有36列齒系。薩斯喀徹溫埃德蒙頓龍的每塊上顎骨有52列齒系,每塊齒骨有44列齒系。
埃德蒙頓龍的脊椎數量,因不同種而有不同變化。帝王埃德蒙頓龍有13節頸椎、18節背椎、9節薦椎、尾椎的數量則不清楚。一個E. annectens標本被認為具有19節背椎、85節尾椎。由於其他鴨嘴龍類的尾椎多為50到70節之間,上述估計值可能過度誇大。埃德蒙頓龍的背部與尾巴脊椎骨具有骨化肌腱,可使背部與尾巴保持僵硬。這些骨化肌腱被認為提供四肢以外的額外支撐脊椎。肩胛骨長而平坦,類似刀狀,與脊椎平行。骨盆包含三個骨頭:長的腸骨、長而薄的坐骨、板狀的恥骨。骨盆的結構允許埃德蒙頓龍以二足方式站立。而9節癒合的薦椎也使骨盆牢固。
分類
埃德蒙頓龍屬於鴨嘴龍科,一群具有鴨嘴型態口鼻部的草食性恐龍,生存於白堊紀晚期。埃德蒙頓龍屬於其中的鴨嘴龍亞科,其頭部缺乏中空頭冠。鴨嘴龍亞科還包含:格里芬龍、慈母龍、短冠龍、納秀畢吐龍、櫛龍、冠長鼻龍、以及原櫛龍。
發現
 埃德蒙頓龍化石
埃德蒙頓龍化石鴨嘴龍科中的鴨龍(Anatosaurus)意為“鴨蜥蜴”。鴨龍曾有五個種,模式種是Anatosaurus annectens,種名意為“連線”。模式種被歸類於埃德蒙頓龍後,鴨龍這個名稱被棄置,而成為埃德蒙頓龍的次異名。而薩斯喀徹溫強龍先是被歸類於薩斯喀徹溫鴨龍(A. saskatchewanensis),後被編入埃德蒙頓龍中,成為了薩斯喀徹溫埃德蒙頓龍。埃德蒙頓鴨龍(A. edmontonensis或A. edmontoni)亦被認為與E. annectens是相同物種。兩個另外的鴨龍種,長頭鴨龍(A. longiceps)及科氏鴨龍(A. copei)被發現與埃德蒙頓龍不同,而被建立為新屬大鴨龍。
埃德蒙頓龍與暴龍生存於相同時期的相同環境,其中一個在丹佛自然科技博物館展覽的成年標本,顯示它的尾巴曾被獸腳類恐龍所咬。尾部的第13到17節脊椎的神經棘具有明顯的牙齒痕跡,其中一個神經棘斷裂,而其他神經棘彎曲,顯示曾經被肉食性動物從後方攻擊過。由於這個部位的高度至少有2.9米,攻擊者應是種體型巨大的動物,而從該化石的發現地區來判斷,該地區的唯一大型肉食性動物是暴龍。骨骼顯示了這個傷口正在痊癒,並有感染的跡象,顯示這頭埃德蒙頓龍在事後應該仍然生存。研究這個標本的肯尼思·卡彭特(Kenneth Carpenter),發現左臀也有個痊癒的骨折跡象,由於已經完全康復,發生時間應較尾巴的傷口早。卡彭特研判這隻埃德蒙頓龍因為左臀的傷口而行動不便,因此成為暴龍的攻擊目標,造成尾巴的新傷口。卡彭特更提出,這隻埃德蒙頓龍成功逃過暴龍的攻擊,或是曾用尾巴抵抗暴龍的嘴部攻擊。另一個發現於南達科塔州的E. annectens標本,身長約7.6米,下頷則具有小型獸腳類恐龍的齒痕。其中部分齒痕已經初步痊癒。Michael Triebold認為曾有小型獸腳類恐龍攻擊埃德蒙頓龍的喉嚨,雖然這隻埃德蒙頓龍在攻擊中存活下來,但不久後因為這個傷口而死亡。
在馬蹄峽谷組中,埃德蒙頓龍生存於接近岸邊的環境,而亞冠龍、櫛龍則生存於較內陸的環境。通常不會在一個地點同時發現埃德蒙頓龍與櫛龍的化石。
社會行為
 埃德蒙頓龍化石
埃德蒙頓龍化石其中,懷俄明州蘭斯組的屍骨層,發現於石灰岩與泥岩中,面積達1平方公里,而埃德蒙頓龍的化石集中於40公頃的地點內。科學家估計該地有10000到25000隻埃德蒙頓龍的化石。科學家也估計這些埃德蒙頓龍聚集於淡水的岸邊,然後死後的屍體被沖積到較深的水域。
在亞伯達省的屍骨層,經常發現艾伯塔龍與蜥鳥盜龍的齒痕。在另一個亞伯達省的化石地點,則在埃德蒙頓龍與櫛龍的化石發現懼龍的齒痕。
埃德蒙頓龍的頭部缺乏骨質頭冠,而鼻孔周圍的骨頭凹陷,科學家推測其鼻孔在生前覆蓋者膨脹的氣囊,可能具有視覺辨識、或發聲的功能。埃德蒙頓龍的體型可分為粗壯、瘦長兩種類型,可能是兩性異形的特徵,但科學家仍無法確定。
由於埃德蒙頓龍的分布廣泛,北至阿拉斯加州,南到科羅拉多州,其中包含北極區,因此有科學家提出埃德蒙頓龍可能具有遷徙過冬的習性。在2008年,Phil R. Bell、Eric Snively提出一份關於恐龍遷徙的研究,認為大部分生存於極區的恐龍可能有過冬的習性,包含:獸腳亞目、蜥腳下目、甲龍下目、棱齒龍類、鴨嘴龍科;而帝王埃德蒙頓龍可能會遷徙達2600公里的路程,每小時大約移動2到10公里。
木乃伊化石
 埃德蒙頓龍化石
埃德蒙頓龍化石這個標本被幾乎完整的保存下來,上附有一些肌肉。皮膚痕跡是因皮膚急速乾透及在泥土上留下形狀而造成的。埃德蒙頓龍的皮膚是有鱗片及皮質的,而肌肉就在皮膚之下。皮膚痕跡只到雙腳的膝蓋,整個大腿都是在皮膚之下。它的外貌就像一隻鴨。在它的頸部、背部及尾巴亦有一些結節。
一個發現於北達科塔州的E. annectens幼年標本,尾巴部份保存了皮膚痕跡,皮膚分布者硬塊與結節、橢圓形的重疊鱗片、有溝紋的鱗片,以及長約9到10厘米的梯形硬塊。
病狀
 埃德蒙頓龍化石
埃德蒙頓龍化石在224塊埃德蒙頓龍的趾骨中,有2.2%發現這個骨軟骨炎等病症,骨軟骨炎是因為軟骨在生長階段未能骨化成骨頭所致。骨軟骨炎的根本原因則未知,目前已有遺傳規律、外傷、攝食強度、血液供應的改變、過量的甲狀腺或荷爾蒙、其他成長因素的缺陷…等理論。
食性
 埃德蒙頓龍頭骨化石
埃德蒙頓龍頭骨化石科學家曾分析埃德蒙頓龍牙齒中的琺瑯質,研究其中的碳與氧同位素比例,以得知埃德蒙頓龍的時性與生理特徵。當動物進食、喝水、呼吸時,輝吸收大自然中的氧與碳,反應在骨頭組織上。不同的外部與內部因素,會影響氧與碳的同位素,例如:該動物的生理狀況、鹽度、氣候。如果骨頭在化石化與成岩的過程中沒有被影響,科學家可以研究其中的同位素,以了解該動物生前的各項因素。以特定植物為食的動物,或是有特定消化過程的動物,身體內會形成特定比例的同位素。形成牙齒琺瑯質的礦物質,使牙齒琺瑯質成為身體中最不受化學變化影響的部分。
在2004年,Kathryn Thomas與Sandra Carlson利用三顆牙齒來研究埃德蒙頓龍的齒系生長狀況,這三顆牙齒髮現於南達科他州科森縣的一個屍骨層,屬於海爾河組,分別是幼年、亞成年、成年的牙齒。他們使用微鑽孔技術,挖出牙齒的內部組織,研究其中的氧同位素比例變化。這三顆牙齒的氧同位素比例變化,似乎與溫暖/乾旱、寒冷/潮濕的氣候變化相關聯。Thomas與Carlson認為這代表者季節變化,也可能是埃德蒙頓龍具有遷徙的習性;但遷徙動物會生存在氣候相近、或有特定食物來源的地區,應會造成平均化的氧同位素比例變化。一顆牙齒的生長,短於0.65年,年輕埃德蒙頓龍的牙齒生長較快。
埃德蒙頓龍的碳同位素數值大,以現代哺乳動物而言,代表它們同時以富含C3的植物(大部分植物),與富含C4的植物(草)為食;但是,白堊紀晚期的富含C4的植物相當稀少。Thomas與Carlson提出可能影響埃德蒙頓龍體內碳同位素的原因,認為這些埃德蒙頓龍最有可能以裸子植物為食,以及西部內陸海道沿岸的耐鹽植物為食;或者是,恐龍與哺乳動物的生理特徵不同,使恐龍的體內形成不同的碳同位素含量。也有可能是以上各因素的綜合結果。
進食方式
 埃德蒙頓龍
埃德蒙頓龍在1984年,大衛·威顯穆沛(David B . Weishampel)首次提出鴨嘴龍類的進食方式研究。威顯穆沛根據鴨嘴龍類的頭部構造,提出下頷可以做出前後移動的動作,而當嘴部咬合時,上頷的骨頭會往外側彎曲。當咬合時,上下頷的牙齒會將嘴中的食物磨碎。這種動作類似哺乳動物的咀嚼動作,但以不同的方式進行。威顯穆沛研究的重要性在於,根據牙齒的磨痕方向,提出顎部的運動方向。雖然其他方向的運動也有可能造成類似的牙齒磨痕。科學家已利用目前技術檢驗這些可能的其他運動方向。
 埃德蒙頓龍
埃德蒙頓龍過去曾有研究宣稱,鴨嘴龍科的一個破碎龍化石曾發現胃石。但該化石是在1900年由巴納姆·布郎(Barnum Brown)發現,被歸類於C. annectens,目前則屬於埃德蒙頓龍的E. annectens。此外,這些胃石其實是化石化過程中,被沖刷、沉積進入的砂礫。
移動方式
埃德蒙頓龍是雙足動物,但亦可以以四足方式行走。雖然前肢較後肢短,但前肢亦有足夠長度,仍適宜行走。前肢的二指有蹄爪,及像圓頂龍的肉墊,可協助分擔重量。後腳有三趾,且都是有蹄爪的。下肢的骨骼結構顯示,腳掌與腳之間是由強壯的肌肉所連線。脊骨在肩膀區段向下彎曲,所以它應該是低姿勢及吃近地面的食物。一個2007年的研究,顯示埃德蒙頓龍能以每小時45公里的時速移動。
