雙殼綱(化石)
正文
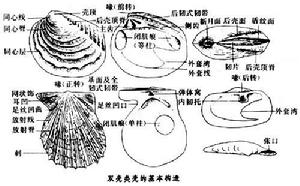
雙殼綱是軟體動物門的1綱,除固著類外一般有兩瓣互為對稱或左右對稱的貝殼。其鰓常呈瓣狀,又稱瓣鰓綱,而它們的頭部退化,有發達呈斧狀的足部,所以也稱無頭綱或斧足綱。本綱動物的殼與腕足動物的相近,但腕足動物殼上具莖孔,背及腹殼大小一般不等也不對稱,而同一個殼呈兩側對稱。這些特徵可與本綱相互區別。雙殼類殼上最早形成的殼尖叫喙(殼嘴)。有些種類由喙向後腹方伸展一條隆脊,叫後殼頂脊,後殼頂脊與後背緣之間的殼面為水管區殼面。在喙與兩瓣鉸緣之間有一較狹的或平或微凹的面,叫基面或鉸合區,還包括外韌帶區。有的在喙前有心形凹陷叫小月面,有的在喙後有長形凹陷,叫盾紋面,有的在鉸緣下前、後端有翼狀的伸出部分,分別稱為前耳與後耳(翼)。前耳下凹入外常有足絲,是用以附著基底的絲狀體。其伸出處叫足絲凹口(右)或凹曲(左)。
殼內外套膜的近外緣部分附著於殼內面上所留下的痕跡叫外套線,有的殼後部外套線向內凹,成為外套灣。閉殼肌附著於殼上的印痕叫閉肌痕。根據閉肌痕的數目和相對大小,分為同柱(前、後兩閉肌痕近於相等)、異柱(後閉肌痕大,前閉肌痕小)和單柱(一個閉肌痕)。在兩殼背部接合處有彈性的韌帶,司殼的開啟。韌帶的附著物主要有3種:基面或韌帶區,是殼外的韌帶附著處;韌片,是殼外喙後韌帶的附著處;彈體窩,是喙下三角形的韌帶附著處。在鉸緣之下有一系列齒與齒窩,司兩瓣的鉸合,合稱齒系。早期較原始種類的一部分,其齒窩及齒數多而弱;較晚期種類的鉸齒漸分異為主齒和側齒。主齒位於喙下,較粗短,與鉸緣呈一定角度相交;側齒遠離喙,多呈片狀,與鉸緣近於平行(見圖)。
 雙殼綱(化石)
雙殼綱(化石)對雙殼綱的起源目前說法不一。60年代曾假設雙殼綱經奧陶紀的多肌痕Babinka屬,由單板綱進化而來。70年代由於對古老的雙殼化石Fordilla屬的研究,以及軟體動物門中建立喙殼綱,並認為通過喙殼綱的Heraulti-pegma屬與雙殼綱的Fordilla屬聯繫起來,而起源於喙殼類。對於中國鄂西下寒武統中發現原始雙殼綱化石的報導,尚有不同意見。
古生物學者的雙殼綱分類依據主要是雙殼綱齒系、貝殼外部形態、肌痕及韌帶等軟體構造遺留下來的痕跡等特徵。但自60年代開始,古生物學者也注重軟體器官鰓、胃和口唇的觸角等並結合鉸合器官綜合進行分類,與貝類學者的分類已趨於一致。
