
概述
 胞間連絲
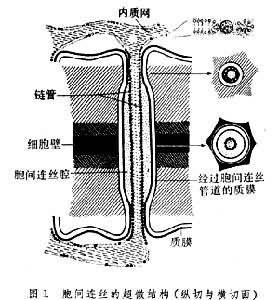
胞間連絲胞間連絲(plasmodesmata)是植物細胞特有的通訊連線。是由穿過細胞壁的質膜圍成的細胞質通道,直徑約20~40nm。因此植物體細胞可看作是一個巨大的合胞體(syncytium)。通道中有一由膜圍成的筒狀結構,稱為連絲小管(desmotubule)。連絲小管由光面內質網特化而成,管的兩端與內質網相連。連絲小管與胞間連絲的質膜內襯之間,填充有一圈細胞質溶質(cytosol)。一些小分子可通過細胞質溶質環在相鄰細胞間傳遞
貫穿兩個相鄰的植物細胞的細胞壁,並連線兩個原生質體的胞質絲。它們使相鄰細胞的原生質連通,是植物物質運輸、信息傳導的特有結構(見彩圖)。這一結構由E.坦格爾於1879年首先在馬錢子胚乳細胞間發現。1882年由德國植物學家、細胞學家E.A.施特拉斯布格命名為“胞間連絲" (plasmodesmen)。
 胞間連絲
胞間連絲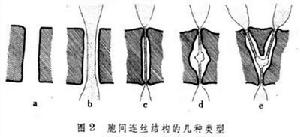 胞間連絲
胞間連絲胞間連絲的結構差異很大,可分5種類型(圖2)。
胞間連絲的主要功能是:①細胞間物質包括小泡的運輸和轉移;②信息、刺激的傳導;③影響細胞的生長、發育和分化。如高等植物頂端分生組織的胞間連絲分布狀況就與分化的控制有關,例如蕨類植物桂皮紫萁胞間連絲在垂周壁與平周壁的分布數目不同就影響了頂端早期葉的發育。此外病毒有時也可經胞間連絲傳播。
 胞間連絲
胞間連絲胞間連絲在功能上與動物細胞間的間隙連線類似,它允許分子量小於800Da的分子通過,在相鄰細胞間起通訊作用。但通過胞間連絲的分子運輸也要受到調節。實驗證明,在胞間連絲正常的情況下,有些低分子量的染料分子卻不能通過。然而某些植物病毒能製造特殊的蛋白質,這種蛋白質同胞間連絲結合後,可使胞間連絲的有效孔徑擴大,使病毒粒子得以通過胞間連絲在植物體內自由播散和感染。
胞間連絲還對細胞分化起一定作用。在高等植物中,頂端分生組織的細胞分化與胞間連絲的分布有著相應的關係。隨著細胞的生長和延長,側壁上的胞間連絲逐漸減少,而橫壁上的卻仍保持很多。植物相鄰細胞間的細胞核可經胞間連絲穿壁。
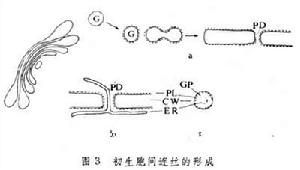
初生胞間連絲是高爾基器小泡融合成細胞板時,因被伸入於其間的內質網膜阻止其融合而形成的(圖3)。
次生胞間連絲是由一些降解酶(果膠酶、半纖維素酶和纖維素酶)的作用使完整的細胞壁穿孔而成。被子植物花粉母細胞間減數分裂早前期出現的次生胞間連絲直徑大,約0.5~1.5微米,特稱細胞融合道。雖然它們能在初生的胞間連絲的位置上形成,但和一般胞間連絲並不完全相同。