
形成特性
組織形成
 植物組織
植物組織植物組織(plant tissue)是由形態結構相似,功能相同的一種或數種類型的細胞組成的結構和
功能單位,也是組成植物器官的基本結構單位。單細胞植物如衣藻等,多細胞群體型植物如團藻、水綿等,它們的每一個細胞都能獨立地完成全部生理功能,因此,這類植物體沒有真正意義上的組織分化。
植物組織的出現是植物進化層次更高的標記。在植物的系統發育過程中,多細胞植物的出現為組織的發生提供了基礎。在多細胞群體型植物向多細胞有機體的進化過程中,群體型個體的細胞間由於所處的位置不同,受到環境的影響也不同。處於不同位置的細胞群間便出現了相異的形態特徵和生理代謝活性與類型的分化。胞間連絲的發生形成,使得相鄰細胞間能夠隨時進行物質、信息和能量的交換,加強了彼此間的聯繫。處於相同位置或同類群的細胞間更加趨於相似或具有同一性。而處於不同位置或不同類群間的細胞也因此而逐漸變得彼此不同。這樣的變化被逐代保留和遺傳下來,成為一種穩定的特性。於是,處於相似或相同位置、有著相似或相同的形態結構和生理功能的細胞群便成了原初類型的組織的共同特徵。
因此,組織是植物在長期適應環境的過程中產生的,其發展和完善也是在適應環境的過程中實現的。植物的進化程度愈高,其體內細胞(群)間的分工愈細,植物體的結構愈複雜,適應性愈強。被子植物是現存植物中高度發達和適應性的植物類群,具有最完善的組織分工,在形態結構和生理功能上表現出高度的統一,適應環境的能力也最強。
在個體發育中,組織的形成是植物體內細胞分裂、生長、分化的結果。組織的形成過程貫穿由受精卵開始、經胚胎階段、直至植株成熟的整個過程。植物體中包含多種組織,它們各有其來源和分工,並有機地組合,協同完成植物的生命活動。
組織屬性
 植物組織
植物組織對於形態結構簡單、沒有器官分化的低等植物而言,組織是其進化發展歷程中的最高形式;
對於形態結構複雜、具有器官分化的高等植物而言,組織是構成複雜有機體的一種結構層次或結構單位。它是處於細胞和組織之間的具有相對獨立性的結構層次。組織以細胞為基本結構單位,同時又是構成器官的基本結構單位。高等的多細胞植物,尤其是種子植物,都是由多種類型的組織構成的。每一類組織的細胞在空間上緊密排列在一起,形態、結構相似,共同完成相同的生理功能。
在高等多細胞植物體內,幾種不同的組織有機配合、緊密聯繫,形成不同的器官(organ),不同的器官之間互相配合,更有效地完成有機體的整個生命活動過程。因此,由細胞到組織,由組織到器官,再到植物體是一個有機整體。
對多細胞植物體而言,細胞、組織或器官都有其相對的獨立性和全息性,在一定的條件下,一個生活細胞、組織或器官的相對獨立的組成部分都可發育成完整植株。組織與組織之間在一定程度上可相互轉化。
植物組織中,細胞的形態結構和它們的生理功能是相適應的。例如,葉肉細胞含有許多葉綠體,執行光合作用、製造有機物質;莖幹具有發達的管狀系統,輸導水分和營養物質,根系的根尖部分表皮細胞外壁凸出,形成毛狀結構,擴大了根的表面,能夠更多地接觸土壤,從中吸收水分和溶於水的無機鹽養分。這些都說明植物組織的形態、結構和功能是高度統一的。
機械組織
機械組織是在植物體內主要起機械支持作用和穩固作用的一種組織。
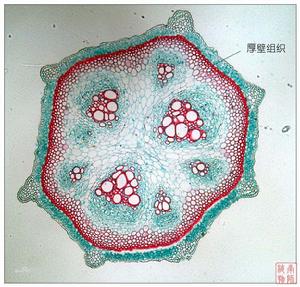
機械組織細胞的特點是其細胞壁均勻或不均勻加厚。根據其細胞的形態、細胞壁加厚程度與加厚方式,可將其分為厚角組織和厚壁組織。
厚角組織
(collenchyma)
厚角組織是一類細胞僅在其角隅處或相毗鄰的細胞間的初生壁顯著增厚的組織(圖2-12左圖)。厚角組織主要出現於雙子葉植物的幼莖、花梗、葉柄以及粗壯的葉脈等的薄壁組織外圍,組成厚角組織的細胞是活細胞,常含葉綠體,有一定的分裂潛能。其細胞壁的局部增厚是伴隨器官的發育而增厚的,因此,厚角組織既有一定的堅韌性,又具有可塑性和延伸性。厚角組織的細胞較長,兩端呈方形、斜形或尖形,彼此重疊連線成束,在橫切面上其細胞腔接近於圓形或橢圓形。
厚壁組織
(sclerenchyma tissue)
厚壁組織是一類細胞壁全面次生增厚、常木質化,細胞腔狹小,成熟細胞一般沒有生活的原生質體的組織。根據其形狀不同又可分為纖維和石細胞。
纖維(fiber) 纖維細胞狹長,兩端尖細,其細胞壁極厚、木質化或少木質化,細胞腔極小,一般沒有原生質體。細胞壁上有少數小的斜縫隙狀紋孔。纖維細胞互相以尖端穿插連線,多成束、成片分布於植物體中,形成植物體內主要的支持結構。根據纖維存在的部位,可將纖維分為韌皮纖維和木纖維,或初生纖維和次生纖維。韌皮纖維主要存在於韌皮部,細胞長、壁厚,其組成主要為纖維素,堅韌性強,可做優質紡織原料;木纖維存在於木質部,細胞短小、壁厚、常木質化,硬度大而韌性差,可做填充料。初生纖維主要指分布於初生韌皮部的纖維,是優質的紡織原料,次生纖維是指分布於次生韌皮部和次生木質部中的纖維,可作為一般工業用纖維。
石細胞(stone cell) 石細胞一般是由薄壁細胞經過細胞壁的強烈增厚並高度木質化(有時也可栓質化或角質化)而來的特化細胞。石細胞的細胞壁堅硬、呈同心層次,分枝紋孔從細胞腔放射狀分出,細胞腔很小,原生質體消失,故具有堅強的支持作用。石細胞形狀多種多樣,或近於短徑、或不規則,最常見的形狀為等直徑、橢圓形、球形、長形、分枝狀或不規則形狀。石細胞通常成群聚生,有時也可單生於植物莖的皮層、韌皮部、髓內,桃[Prunus persica(L.) Batsch]、杏(Prunus armeniacaL.)等的果皮,和蠶豆(Vicia fabaL.)等的種皮中也較常見。茶(Camellia sinensisO.Ktze.)等的葉肉、梨(Pyrus bretschneideriRehd.)等的果肉中分布的石細胞形態、數量是鑑別其品質的依據。石細胞的形態特徵有時也作為區分物種的參考依據。
輸導組織
(conducting tissue)
輸導組織是植物體內長距離運輸物質的組織,其細胞長管狀、相互貫通成為統一的整體。根據輸導組織的結構和所運輸的物質不同,可將其分為運輸水分和無機鹽類的導管與管胞,以及運輸有機同化物的篩管與篩胞兩大類。
(1)導管(vessel)
導管普遍存在於被子植物的木質部,向上運輸根從土壤中吸收的水分和無機鹽。它們是由許多管狀的、細胞壁木質化的死細胞縱向連線成的一種輸導組織。組成導管的每一個細胞稱為導管分子(vessel element)。
在導管分子的發育初期,其細胞內含有豐富的微管、內質網、高爾基體等細胞器。隨著細胞的生長、液泡的分化和繼後的細胞程式性死亡過程,導管分子的側壁形成各種紋飾的次生加厚,端壁逐漸解體消失,形成不同形式的穿孔(單穿孔或數個小孔組成的復穿孔)。具有穿孔的端壁稱為穿孔板(perforation plate)。穿孔的形成及原生質體消失使導管成為中空的連續長管,有利於水分及無機鹽的縱向運輸。導管還可通過側壁上的紋孔或未增厚的部分與毗鄰的細胞進行橫向運輸。根據導管的發育先後和側壁木質化增厚方式,可將其分為環紋導管、螺紋導管、梯紋導管、網紋導管和孔紋導管五種類型。
環紋導管(annular vessel)和螺紋導管(spiral vessel) 這類導管是在器官的初生生長早期形成的,位於初生木質部中的原生木質部,其導管分子細長而腔小(尤其是環紋導管),且其側壁分別呈環狀或螺鏇狀木質化加厚,輸水能力弱,有時同一條導管的不同部分可出現環紋與螺紋增厚。由於其增厚的部分不多,未增厚的管壁部分仍可適應於器官的生長而伸延,但易被拉斷。
梯紋導管(scalariform vessel)、網紋導管(reticulated vessel)和孔紋導管(pitted vessel) 這類導管是在器官的初生生長中後期和次生生長過程中形成的,位於初生木質部中的後生木質部和次生木質部,其導管分子短粗而腔大,輸水效率高(尤其是孔紋導管)。梯紋導管和網紋導管的側壁分別呈梯狀和網狀增厚,孔紋導管的側壁則大部分木質化增厚,未加厚的部分則形成紋孔。
(2)管胞 (tracheid)
管胞是絕大部分蕨類植物和裸子植物的唯一輸水組織,多數被子植物中,管胞和導管可同時存在於木質部中。管胞長梭狀,兩端尖斜不穿孔,橫切面呈三角形、方形或多角形。管胞次生壁的木質化和增厚方式與導管相似,在側壁上也呈現環紋、螺紋、梯紋和孔紋等多種方式的加厚紋飾。
(3)篩管(sieve tube)和伴胞(companian cell)
篩管存在於被子植物的韌皮部中,是由若干不具細胞核的管狀活細胞(即篩管分子,sieve element)縱向連線而成的、運輸有機物質的輸導組織。篩管分子的細胞壁不次生加厚,端壁(稱為篩板,sieve plate)上存在著成群的小孔——篩孔(sieve pore),具有篩孔的凹陷區域稱為篩域(sieve area),只有一個篩域的端壁叫單篩板,具有多個篩域的端壁叫復篩板。穿過篩孔連線相鄰兩個篩管分子的原生質呈束狀,稱為聯絡索(connecting strand)。聯絡索通過篩孔彼此相連,使縱接的篩管分子相互貫通,形成運輸同化產物的通道。
在每個篩管的旁邊通常有一個或多個伴胞(companion cell)。伴胞與篩管在發育上具有同源性(由同一母細胞分裂而來),在功能上伴胞從屬於篩管,協助和保證篩管的活性與運輸功能。
(4)篩胞(sieve cell)
篩胞是蕨類植物和裸子植物體內主要承擔輸導有機物的細胞。篩胞通常細長,末端尖斜,細胞壁上可有不甚特化的篩域出現,篩孔細小、一般不形成篩板結構。許多篩胞的斜壁或側壁相接而縱向疊生。運輸有機物質的效率比篩管差得多,是比較原始的運輸有機物質的組織。
導管和篩管是被子植物體內物質輸導的重要組織,但也是病菌感染、傳播擴散的主要通道。如土壤中的枯萎病菌入侵根部後,其菌絲可隨導管到達地上部分的莖和葉,某些病毒可借昆蟲刺吸取食而進入篩管,引起植株發害。因此,研究輸導組織的特性,有利於合理施用內吸傳導型農藥,有效防治病、蟲、草害。
5)異細胞
異細胞是單獨分散於其它各種組織中的含特殊分泌物的細胞。異細胞由薄壁細胞特化而來,其細胞壁稍厚、細胞體積較大,細胞中的內含物種類很多,有分類學價值,如內含揮髮油、樹脂、樹膠、單寧、蛋白質或粘液等次生物質(含單寧異細胞、含晶異細胞等)。
異細胞的研究對於了解植物的生理意義和生化調節機制有著重要的意義。
複合組織
(compound tissue)
複合組織是指由發育上或功能上密切相關的幾種簡單組織組成的、共同執行特定生理功能的、相對獨立的結構。複合組織位於植物體的特定部位。如周皮、樹皮、木質部、韌皮部、維管束、分泌結構等都是由多種組織複合而成。
1)周皮(periderm)
周皮由木栓層、木栓形成層和栓內層共同構成。從組織分類來看,木栓層屬於次生保護組織,木栓形成層屬於次生分生組織,而栓內層屬於次生薄壁組織,因此,周皮是由三種不同組織組成的複合組織。
2)木質部(xylem)和韌皮部(phloem)
木質部和韌皮部是植物體內主要起輸導和支持作用的組織。木質部一般包括導管(多數蕨類植物及裸子植物無導管)、管胞、木薄壁組織和木纖維等;韌皮部包括篩管、伴胞(蕨類植物及裸子植物僅為篩胞,無篩管、伴胞)、韌皮薄壁組織和韌皮纖維等。木質部和韌皮部的組成分子包含輸導組織、薄壁組織和機械組織等幾種組織,因而被稱為複合組織。由於木質部或韌皮部的主要組成分子都是管狀結構,因此,常將木質部和韌皮部或者將其中之一稱為維管組織(vascular tissue)。維管組織的形成,在植物系統進化過程中,對於適應陸生生活有著重要意義。從蕨類植物開始,它們體內已有維管組織的分化出現。種子植物體內的維管組織則更為發達進化。通常將蕨類植物和種子植物總稱為維管植物。
3)維管束(vascular bundle)
維管束存在於蕨類植物和種子植物中,是由原形成層分化而來、由木質部和韌皮部組成的束狀結構。在不同種類的植物或不同的器官內,原形成層分化成木質部和韌皮部的情況不同,也就形成不同類型的維管束。根據維管束內束中形成層的有無或能否增粗生長,可將其分為有限維管束和無限維管束兩大類型。
(1)有限維管束(closed bundle)
有限維管束是其組成上僅有木質部和韌皮部、沒有束中形成層的維管束。這類維管束一經形成便不再增粗,如大多數單子葉植物中的維管束等。
(2)無限維管束(open bundle)
無限維管束是指組成上含有除木質部和韌皮部外,在二者之間還保留有束中形成層的維管束。這類維管束以後能通過形成層的分裂活動,不斷產生次生韌皮部和次生木質部,如裸子植物和許多雙子葉植物的維管束。
4)分泌結構(secretory structure)
植物體中凡能產生分泌物質(如糖類、揮髮油、有機酸、乳汁、蜜汁、單寧、樹脂、生物鹼、抗生素等)的有關細胞或特化的細胞組合稱為分泌結構。分泌結構一般由分泌細胞和其他薄壁細胞所組成。
通常根據分泌結構的存在部位和分泌物的溢排情況,將其劃分為外分泌結構和內分泌結構兩類。
(1)外分泌結構(external secretory structure)
外分泌結構分布在植株的外表,其分泌物排於體外的分泌結構,如腺毛、腺鱗、鹽腺、蜜腺和排水器等。
腺毛和腺鱗 腺毛(glandular hair)一般具有頭部和柄部兩部分,頭部由單個或多個產生分泌物的細胞組成,柄部是由不具分泌功能的薄壁細胞組成,著生於表皮上。
蜜腺 蜜腺(nectary)是能分泌蜜汁的多細胞腺體結構,它們由表皮或表皮及其內層細胞共同形成,即由保護組織和分泌細胞構成。根據蜜腺在植物體上的分布位置,可將蜜腺分為花蜜腺和花外蜜腺兩類。
鹽腺 鹽腺(salt gland)是將過多的鹽分以鹽溶液狀態排出植物體外的外分泌結構。
排水器 排水器(hydathode)是植物將體內多餘的水分直接排出體外的結構,常分布於植物的葉尖和葉緣。從排水器排水過程稱為吐水(guttation)。在溫濕的夜間或清晨,常在葉尖或葉緣出現水滴,就是經排水器泌出的水液。吐水現象往往可作為根系正常生長活動的一種標誌。
(2)內分泌結構(internal secretory structure)
內分泌結構是將分泌物貯存於植物體內的分泌結構。它們常存在於基本組織內,常見的有分泌囊(腔)、分泌道和乳汁管。
分泌細胞團 分泌細胞團(secretory cell nodule)一般由數層扁平的鞘細胞包裹著一團分泌細胞構成,其中央無腔隙。
分泌囊(腔) 分泌囊(secretory cavity)是由外側1~2層扁平的鞘細胞包裹著1層長方形的上皮細胞(第一層分泌細胞)及其內的分泌腔共同構成。分泌腔是植物體內多細胞構成的儲藏分泌物的腔室結構。根據腔室形成的方式可分為溶生分泌腔和裂生分泌腔兩種類型。溶生(lysigenous)分泌腔較多,最初有一群具有分泌能力的分泌細胞,漸漸地細胞內的分泌物質增多,最後細胞解體,形成溶生的腔室,原來細胞中的分泌物貯積在腔中,形成分泌腔,如橘(Citrus reticulataBlanco)的果皮和葉,棉的莖、葉、子葉中都有這種類型的分泌腔。裂生(schizogenous)分泌腔是構成分泌腔的細胞間隙擴大成的腔室,周圍1至多層分泌細胞將分泌物排入腔室中。
分泌道 分泌道(secretory canal)為管狀的內分泌結構,管內貯存分泌物質。分泌道也有溶生和裂生的方式,但多為裂生而形成。如松柏類植物的樹脂道即是分泌細胞的胞間層溶解,細胞相互分開而形成的長形細胞間隙,完整的分泌細胞環生於分泌道周圍,由這些分泌細胞分泌的樹脂貯存於分泌道中。樹脂的產生,增強了木材的耐腐性。漆樹中有裂生的分泌道稱為漆汁道,其中貯有漆汁。樹脂和漆汁都是重要的工業原料,經濟價值很高。
乳汁管 乳汁管(laticifer)是能分泌乳汁的管狀結構。按其形態發生特點可分為無節乳汁管和有節乳汁管。如大戟屬(Euphorbia)和萵苣(Lactuca sativaL.)等的乳汁管。
分生組織(meristem)
簡介
分生組織存在於高等植物體內的特定部位,是一類可連續性或周期性分裂產生新細胞的組織。分生組織的細胞經過分裂、生長、分化而形成其他各類組織,直接關係到植物的生長和發育。
分生組織的細胞排列緊密、一般無細胞間隙; 細胞壁薄、主要由果膠和纖維素構成;細胞核相對較大,細胞質濃、細胞器豐富,但—般沒有液泡和質體的分化。
根據分生組織的發育來源和在植物體中的分布位置,可將分生組織分為不同類型。
1.根據分生組織的來源、發育程度和性質,將其分為原生分生組織、初生分生組織和次生分生組織。
原分生組織
(promeristem)
原分生組織包括胚和成熟植株的莖尖或根尖的分生組織先端的原始細胞。原分生組織的細胞體積較小、近於正方體、細胞核相對較大、細胞質濃、細胞器豐富,有很強的持續分裂或潛在分裂能力。是產生其它組織的最初來源。
| | |
初生分生組織
(primary meristem)
初生分生組織位於根端和莖端的原分生組織的後方,是原分生組織細胞分裂後經有限生長或衍生而來的組織。初生分生組織的細胞分裂能力仍較強,部分細胞初步分化為原表皮、基本分生組織和原形成層。原表皮(protoderm)位於最外周,主要進行徑向分裂;基本分生組織(ground meristem)位於原表皮之內,所占比例最大,可進行各個方向的分裂,以增加分生組織的體積;原形成層(procambium)位於基本分生組織中的特定部位,其細胞扁而長,是分化產生成熟組織的基礎)。
次生分生組織
(secondary meristem)
次生分生組織是由某些成熟組織經脫分化、重新恢復分裂能力而來的組織。次生分生組織的細胞或扁長或短軸型的扁多角形,細胞呈不同程度的液泡化。次生分生組織包括木栓形成層(phellogen)和維管形成層(vascular meristem)(尤其是束間形成層),主要分布於根莖器官的內側,並與其長軸相平行,與根莖的逐年增粗直接相關
2.根據分生組織在植物體中的存在位置,將其分為頂端分生組織、側生分生組織和居間分生組織
頂端分生組織
(apical meristem)
頂端分生組織位於根和莖頂端的分生區部位(圖2-2),其細胞小、排列緊密、近於方形,能較長期地保持旺盛的分裂能力。頂端分生組織分裂產生的細胞,一部分繼續保持分裂能力,一部分逐漸分化、形成各種有關的成熟組織。頂端分生組織是根、莖、腋芽和幼葉生長的基礎。當有花植物發育到一定階段,莖的頂端分生組織可轉化為產生花或花序的分生組織。
側生分生組織
(1ateral meristem)
側生分生組織是一類分布於植物體內、平行於所在器官的表面,且與所在器官的增粗有關的次生分生組織(圖2-4)。側生分生組織包括維管形成層和木栓形成層,其細胞特徵與次生分生組織相同。在多年生植物體內,維管形成層可逐年活動,產生新的細胞,位於維管形成層外側的細胞則分化為次生韌皮部,位於其內側的分化為次生木質部;木栓形成層的分裂活動,向外形成木栓層,向內形成栓內層,復蓋於老根和老莖的外周。側生分生組織活動的結果,使裸子植物和多數雙子葉植物的根和莖得以增粗。
居間分生組織
(intercalary meristem)
居間分生組織位於莖、葉、子房柄、花梗、花序軸等器官節段的基部或其成熟組織之間,其細胞的分裂僅局限於一定時空,便轉變為成熟組織。居間分生組織的細胞核大,細胞質濃,有一定程度的液泡化,主要進行橫向分裂,使器官縱向伸長。禾本科植物莖的每個節間的基部都具有居間分生組織,如水稻(
L.)、小麥(
L.)等拔節、抽穗,竹類植物的雨後春筍,韭菜(
Rottler)、蔥(
L.)葉子刈割後的快速生長,以及花生(
L.)子房的“入土結實”等,都與居間分生組織的活動有關。
新近的研究指出:分生組織的活性是多個 基因相互作用的結果。如STM(shoot meristemless)等是決定擬南芥 苗端分生組織特徵的關鍵基因。
不同分類標準的各類分生組織間的關係
從組織發生的來源和性質分析:頂端分生組織包括原分生組織和初生分生組織。因此,頂端分生組織既具有原分生組織的主要特徵即較強的分裂能力,又具有初生分生組織的特性:一方面能夠保持細胞分裂能力,另一方面向成熟組織分化。居間分生組織是由莖尖頂端分生組織衍生而遺留在某些器官局部區域的分生組織,屬於初生分生組織。側生分生組織的細胞是在初生分生組織分裂的細胞經生長、分化、成熟基礎上,由成熟組織經脫分化、恢復分裂而來。因此,側生分生組織屬於次生分生組織。
