
分子機理
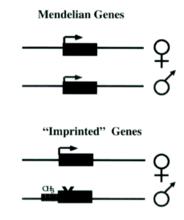 甲基化——一種基因組印跡的原理示意圖
甲基化——一種基因組印跡的原理示意圖研究發現,基因組印跡的分子機理與印跡基因DNA中胞嘧啶甲基化尤其是CpG島的甲基化密切相關,胞嘧啶甲基化是DNA的一種共價修飾。另外還有特殊的染色質結構和反義轉錄產物等可能都是基因印跡產生和維持的重要因素。
基因印跡中,卵子和精子中對同一基因的不同程度的甲基化,幾乎所有的印跡基因都有一些序列成份在兩個不同親本來源的等位基因中僅有一方是甲基化。這些序列被稱為“差異甲基化區域“(Differentially Methylated Regions)。
它指基因組在傳遞信息的過程中,對基因或DNA片段打下標記或烙印的過程。
印跡基因特點
分布的集群性
最大的印跡基因群集區為X染色體本身,而最大的印跡基因群集區之一位於小鼠7號染色體末端和人類11p15.5處。
一個研究較多的印跡基因群集區位於人類15號染色體上,由於該區與普達-威利綜合症和亞斯伯格症候群兩種人類遺傳疾病有關,因此人們對這一群集區很感興趣,目前已發現在此群集區至少包括ZNF127、VBE3A、5NRPN、 PAR-SN、PAR5、PAR1、NDN (necdin)、IPW、GABRG3、 GABRB3、GABRA5和FNZ127等12個印跡基因。
DNA複製的不同步性
細胞周期中的複製時間常與基因表達水平有關,早複製的基因有活性,但印跡基因並不一定遵循此規則。
基因表達的時空特異性
時間特異性:如小鼠胚胎中的Igf2和Mash2開始為雙等位基因表達,但在胚胎形成的後期則表現為母源等位基因表達。人類印跡基因中Igf2基因的表達也有時間改變。
空間特異性:如小鼠Ins-2基因僅在小鼠胚胎的胚外組織中印跡,但在胰島細胞中雙等位基因均表達;在人類,KvLQT1基因在大多數組織中為母源表達,但在心臟中無印跡。
基因的保守性
人們對小鼠和人類基因組印跡基因研究發現人類和小鼠的印跡基因既有相似又有不同點。至今還沒有發現真獸亞綱以外的動物有基因組印跡現象的報導,但有袋類動物的X染色體上有印跡現象,它的失活不是隨機的,在胚胎和胚外組織中均為父源X染色體優先失活。
印跡基因編碼RNA
人們發現很多印跡基因根本不編碼蛋白質,只編碼RNA。這類編碼RNA的基因具有一些共同特點:這些基因的第一個和最後一個外顯子均大,中間含有多個小的外顯子;在真獸亞綱動物中兩個基因編碼的RNA一級結構均很保守,雖然那些保守的核苷分布並不完全一致,但在每個基因中其出現具有周期性。根據潛在的基-環結構上互補鹼基變化方式推測出RNA的二級結構其保守性更加顯著。這特點表明印跡基因編碼的RNA在進化選擇中有一定的生物學活性。
用例
印跡的抑癌基因的雜合性丟失(LOH)、UPD或突變失活可能會導致某個抑癌基因的唯一有功能拷貝的丟失或不表達。
印跡的癌基因的LOI或UPD則可能導致雙等位基因表達,表達量成倍增加。
印跡控制中心(指理論上可能會存在的某些對印跡現象有關鍵性影響的基因或順式元件)的突變性失活, 可能會導致某個染色體印跡區域的多個印跡的癌基因的不正常表達。
生物學意義
 印跡基因對進化的研究有重要意義
印跡基因對進化的研究有重要意義印跡基因在生長發育中尤其是胎兒和胎盤的生長發育中有重要作用,它還與細胞增殖有關,正常基因組印跡模式改變會引起一系列人類遺傳性疾病,包括神經和精神發育異常的遺傳性疾病以及兒童和成人的一些腫瘤。
印跡基因還能幫助人們認識生物進化的本質。基因印跡在進化論意義上的優勢可能就是有效地防止了單性生殖, 維持了遺傳多樣性, 但也增加了隱性突變轉為顯性的危險性。
