
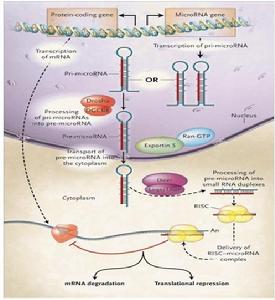
 microRNA
microRNA形式
1. pre-miRNA
約70bp含microRNA莖環結構的pre-miRNA。
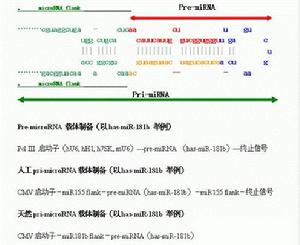
製備方式:化學合成、生物轉錄合成、pre-miRNA質粒表達載體、pre-miRNA病毒。
2. pri-miRNA
天然pri-miRNA
從染色體基因文庫中調取300bp-1000bp完整的microRNA基因,克隆到質粒載體(普通載體或病毒載體),以強大的CMV啟動子操縱該300bp-1000bp microRNA。
人工pri-miRNA
選擇一個完整的microRNA基因,克隆到質粒載體(普通載體或病毒載體)。以人工合成的約70bp含miRNA莖環結構的目標pre-RNA替代原pre-RNA,並以pol Ⅱ/pol III 啟動子操縱該microRNA結構單元。
作用方式
 microRNA的形式
microRNA的形式特異性
研究表明MiRNAs在物種間具有高度的保守性、時序性和組織特異性——在特定的時間、組織才會表達。
細胞特異性或組織特異性是miRNA表達的主要特點,又如擬南芥中的miR-171僅在其花序中高水平表達,在某些組織低水平表達,在莖、葉等組織中卻無任何表達的跡象;20-24h的果蠅胚胎提取物中可發現miR-12,卻找不到miR3-miR6,在成年果蠅中表達的miR-1和let-7也無法在果蠅胚胎中表達,這同時體現了miRNA的又一特點——基因表達時序性。MiRNA表達的時序性和組織特異性暗示miRNA的分布可能決定組織和細胞的功能特異性,也可能參與了複雜的基因調控,對組織的發育起重要作用。
siRNA是RNAi途徑的主要作用物,miRNA和siRNA很容易混淆,他們有許多共同點也有許多不同點。為了能夠清楚地讓讀者弄清兩者之間的差異之處,筆者特別將它們之間的差別劃分入三個大的階段:起源階段、成熟階段和功能階段(即調節基因表達的作用階段)。
在描述兩者的差異之前,有必要先說一說它們的共同點:
1. MiRNA和siRNA都是由22個左右的核苷組成;
2. 它們都是Dicer酶的產物;
3. 它們在起干擾、調節作用時都會和RISC複合體結合;
4. 它們都可以在轉錄後和翻譯水平干擾以抑制靶標基因的翻譯;
差異
兩者之間的主要差異:
起源階段
SiRNA:通常是外源的,如病毒感染和人工插入的dsRNA被剪下後產生外源基因進入細胞(註:病毒入侵,或者是自身合成RNA中出現錯誤,細胞內就會產生雙鏈RNA,來阻止這些異常基因的表達)。
MiRNA:是內源性的,是一種非編碼的RNA;由miRNA基因表達出最初的pri-miRNA分子。
成熟過程
SiRNA:直接來源是長鏈的dsRNA(通常為外源);經過Dicer酶*切割形成雙鏈siRNA,而且每個前體daRNA能夠被切割成不定數量的siRNA片段。
MiRNA:在細胞核中轉錄的較大的pri-miRNA經由Drosha(一種RNAse Ⅲ酶)和Pasha(含有雙鏈RNA結合區域)加工成為單鏈pre-miRNA;接著,髮夾狀、部分互補的pre-miRNA在細胞質中被Dicer*(一種RNAse Ⅲ酶)酶切割形成miRNA;在生物體中的表達具有時序性、保守性和組織特異性。
功能階段
siRNA:它與RISC*(RNA誘導的沉默複合物,使用的AGO蛋白家族的成分為AGO2)結合,以RNAi途逕行使功能,即通過與序列互補的靶標mRNA完全結合(與編碼區結合),從而降解mRNA以達到抑制蛋白質翻譯的目的;它通常用於沉默外源病毒、轉座子活性。
MiRNA:它和RISC形成複合體(利用的AGO蛋白家族成員為AGO1)後與靶標mRNA通常發生不完全結合,並且結合的位點是mRNA的非編碼區的3’端;它不會降解靶標mRNA,而只是阻止mRNA的翻譯; miRNA能夠調節與生長發育有關的基因。
註:RISC, RNA誘導的沉默複合物(RNA-induced silencing complex; RISC)的組裝是在RNAi和miRNA通路中最為複雜的過程。新的研究表明,與siRNA和miRNA結合的RISC複合物並不完全相同其中的AGO蛋白質有AGO1和AGO2之分。剛產生的siRNAs和miRNAs都是雙鏈結構,這種雙鏈結構需要解螺鏇才能被組裝到RISC中發揮作用。組裝後的複合物分別稱為siRISC和miRISC。從dsRNA引發RNAi的發生大致劃分為三個階段,即啟動、剪下和擴增。
Dicer酶:新的研究表明siRNA成熟需要Dicer-2和R2D2蛋白,而miRNA則依賴Dicer1和它的伴侶loqs蛋白。
選擇方法
1.pre-miRNA是最早採用的microRNA,擁有大量的成功報導。化學合成的pre-miRNA製備快捷,可以標記螢光等追蹤。缺點是製備的RNA穩定性較差,而且,由於製備的microRNA較長,合成時RNA的錯誤難以避免(當合成RNA超過60bp時,出現一個鹼基錯誤率約30%),轉錄製備的pre-miRNA穩定性較好,但無flank結構,很少使用。質粒載體形式的pre-miRNA也因為發現microRNA的flank對於microRNA功能和測定非常重要,現已逐漸被pri-miRNA取代。
2.人工pri-miRNA效果較pre-miRNA好,也有足夠多的成功使用的經驗報導,但這種人工microRNA採用固定的mir155 flank,製備的microRNA功能不及天然原始microRNA,故也逐漸較少使用。
3.原始天然pri-miRNA克隆自天然文庫的microRNA由於保留了每個microRNA獨自的原始天然雙臂結構,效果更好,是近來被首選方法。
4.pre-miRNA或pri-miRNA腺病毒:可以轉染大多數絕細胞,產品質量可靠。缺點是腺病毒有一定的毒性,以往實驗腺病毒毒性並未引起重視,但我們發現在microRNA實驗中顯得比較突出。另外,製備周期長,費用高也是不容忽視的缺點。
5.慢病毒microRNA:目前最好的microRNA實驗產品,缺點是慢病毒自身的不足,即製備lenti-virus時,病毒的質量批間差異較大。
AAV、或retrovirus microRNA:與腺病毒和lenti-virus microRNA一樣,具有病毒產品的優勢與缺點。
microRNA的檢測
越來越多的研究成果表明,miRNA將在疾病,特別是癌症的診斷和治療中起到積極的深遠作用。與蛋白編碼基因表達譜相比,用miRNA表達譜對癌症進行分型更加精確可靠。此外,在常規收集的、福馬林固定、石蠟包埋的臨床組織標本中,miRNAs都表現出較高的穩定性,更進一步預示其作為診斷性分子標記的巨大潛能。因此,研究特定細胞microRNA的表達圖譜、篩選未知miRNA、探究每種miRNA的特定功能成為當前的研究熱點。
目前常見的miRNA檢測方法包括NorthernBlot、微陣列晶片、微球、實時螢光定量PCR等技術,然而由於以下原因而各具缺陷。首先,由於miRNA本身鹼基數過少,使得探針鹼基序列的選擇受限。並且不同microRNA具有不同的最適雜交溫度,在相同的雜交條件下,往往會引起很大程度的錯配雜交,從而限制了對於相似序列microRNA的高效識別。其次,由於miRNA存在序列的高度相似性,所以檢測的特異性往往難以保證。
目前市場占主導地位的商品是Invitrogen公司(原ABI公司)開發的實時螢光定量PCR方法(TaqManmicroRNA檢測方法),該方法雖然在一定程度上克服了上述缺陷,但是仍具有高成本等缺點,限制了其廣泛套用。
北京賽諾亞生物技術有限責任公司(www.sirnoa.com)自主開發出了方便實用的高通量miRNA檢測方法:等溫實時螢光定量miRNA檢測[1]。該技術具有高特異、高靈敏度、低成本等特點。而且,樣品不需要經過特殊處理,可直接提取RNA後用來檢測,檢測靈敏度高達1000拷貝數;相對於市場上占主流的InvitrogenTaqManmiRNA檢測方法,該方法在成本和特異性等方面具有明顯優勢。
