概述
 Notch信號途徑
Notch信號途徑Notch基因最早發現於果蠅,部分功能缺失導致翅緣缺刻(notches)。在胚胎髮育中,當上皮組織的前體細胞中分化出神經元細胞後,其細胞表面Notch配體Delta與相鄰細胞膜上的Notch結合,啟動信號途徑,防止其它細胞發生同樣的分化,這種現象叫作側向抑制(lateralinhibition)。Notch突變的半合子[9]或純合子在胚胎期死亡,其胚胎中神經組織取代了上皮組織從而使神經組織異常豐富。
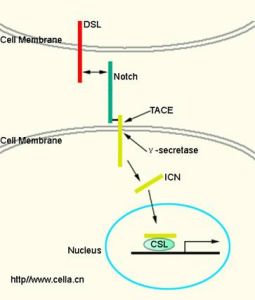
Notch信號途徑由Notch、Notch配體(DSL蛋白)和CSL(一類DNA結合蛋白)等組成。Notch及其配體均為單次跨膜蛋白,當配體(如Delta)和相鄰細胞的Notch結合後,Notch被蛋白酶體切割,釋放出具有核定位信號的胞質區ICN(intracellulardomainofNotch),進入細胞核與CLS結合,調節基因表達。可概括為:Delta→Notch→酶切→ICN→進入細胞核→CLS-ICN複合體→基因轉錄。
詳細介紹
Notch:為分子量約300KD的蛋白質,果蠅只有1個Notch基因,人類4個(Notch1-4)。Notch的胞外區是結合配體的區域,具有不同數量的EGF樣重複序列(EGF-R)和3個lnr(Lin/Notchrepeats)。胞內區由RAM(RBP-Jkappaassociatedmolecular)結構域、6個錨蛋白(cdc10/ankyrin,ANK)重複序列、2個核定位信號[10](NLS)和PEST結構域。RAM結構域是與CSL結合的區域,PEST結構域與Notch的降解有關。Notch蛋白要經過三次切割,第一次在高爾基體內被furin切割為2個片斷,轉運到細胞膜形成異二聚體。當配體結合到胞外區,Notch蛋白又發生兩次斷裂,先是被腫瘤壞死因子-α-轉化酶(TNF-α-convertingenzyme,TACE)切割,然後被γ-促分泌酶(γ-secretase)切割,後者需要早老蛋白(presenilin,PS)參與。酶切以後釋放Notch胞內區ICN,進入細胞核發揮生物學作用。
Notch配體:在果蠅中Notch個配體為Delta和Serrate,線蟲為Lag-2,取首寫字母,Notch的配體又被稱為DSL蛋白(在哺乳動物中叫做Jagged),都是單次跨膜糖蛋白,其胞外區含有數量不等的EGF樣重複區,N端有一個結合Notch體必需的DSL基序。
CSL:為轉錄因子,在哺乳動物中叫做CBF1,在果蠅中叫做SuppressorofHairless,線上蟲中叫做Lag-1,故名。CSL能識別並結合特定的DNA序列(GTGGGAA),這個序列位於Notch誘導基因的啟動子上。ICN不存在時,CSL為轉錄抑制因子。當結合ICN時,CSL能誘導相關基因的表達。Notch信號的靶基因多為鹼性螺旋-環-螺旋類轉錄因子(basichelix-loop-helix,bHLH),它們又調節其它與細胞分化直接相關的基因的轉錄。如哺乳動物中的HES(hairy/enhancerofsplit)、果蠅中的E(spl)(enhancerofsplit)及非洲爪蟾中的XHey-1等。
