
定義
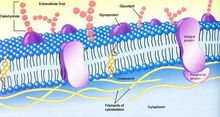 質膜
質膜圍繞各種細胞器的膜,稱為細胞內膜。質膜和內膜在起源、結構和化學組成的等方面具有相似性,故總稱為生物膜(biomembrane)。生物膜是細胞進行生命活動的重要物質基礎,細胞的能量轉換、蛋白質合成、物質運輸、信息傳遞、細胞運動等活動都與膜的作用有密切的關係。
真核細胞除了具有質膜、核膜外,發達的細胞內膜形成了許多功能區隔。由膜圍成的各種細胞器,如內質網、高爾基體、線粒體、葉綠體、溶酶體等。在結構上形成了一個連續的體系,稱為內膜系統( endomembrane system )。內膜系統將細胞質分隔成不同的區域,即所謂的區隔化( compartmentalization )。區隔化是細胞的高等性狀,它不僅使細胞內表面積增加了數十倍,各種生化反應能夠有條不紊地進行,而且細胞代謝能力也比原核細胞大為提高。
質膜表面寡糖鏈形成細胞外被(cell coat)或糖萼(glycocalyx);質膜下的表層溶膠中具有細胞骨架成分組成的網路結構,除對質膜有支持作用外,還與維持質膜的功能有關,所以這部分細胞骨架又稱為膜骨架。細胞外被、質膜和表層胞質溶膠構成細胞表面。
研究歷史
1. E. Overton 1895 發現凡是溶於脂肪的物質很容易透過植物的細胞膜,而不溶於脂肪的物質不易透過細胞膜,因此推測細胞膜由連續的脂類物質組成。
2. E. Gorter & F. Grendel 1925 用有機溶劑提取了人類紅細胞質膜的脂類成分,將其鋪展在水面,測出膜脂展開的面積二倍於細胞表面積,因而推測細胞膜由雙層脂分子組成。
3. J. Danielli & H. Davson 1935 發現質膜的表面張力比油-水界面的張力低得多,推測膜中含有蛋白質,從而提出了”蛋白質-脂類-蛋白質”的三明治模型。認為質膜由雙層脂類分子及其內外表面附著的蛋白質構成的。1959年在上述基礎上提出了修正模型,認為膜上還具有貫穿脂雙層的蛋白質通道,供親水物質通過。
4. J. D. Robertson 1959 用超薄切片技術獲得了清晰的細胞膜照片,顯示暗-明-暗三層結構,厚約7.5nm。這就是所謂的“單位膜”模型。它由厚約3.5nm的雙層脂分子和內外表面各厚約2nm的蛋白質構成。單位膜模型的不足之處在於把膜的動態結構描寫成靜止的不變的。
5. S. J. Singer & G. Nicolson 1972根據免疫螢光技術、冰凍蝕刻技術的研究結果,在”單位膜”模型的基礎上提出”流動鑲嵌模型”。強調膜的流動性和膜蛋白分布的不對稱性,
但並不能說明具有流動性的質膜在變化過程中怎樣保持膜的相對完整性和穩定性。
6. Wallach 1975 提出“晶格鑲嵌模型”。認為,膜蛋白對脂類分子的運動有限制作用,鑲嵌蛋白和其周圍的脂類分子形成膜中晶態部分,而具有流動性的脂類呈小片的點狀分布。因此,脂類的流動性是局部的,並非整個脂類雙分子都在進行流動,這就比較合理地解釋了生物膜既具有流動性又可以保持相對完整性和穩定性的原因
。
7.Jain &White 1977 提出“板塊鑲嵌模型”。認為在流動的脂雙分子層中存在許多大小不同、剛性較大的能獨立移動的脂類板塊,這些有序結構板塊間存在流動的脂類 ,二者處於連貫的動態平衡
。
6. 1988年,提出“脂筏模型”。即脂雙分子層並不是一個完全均勻的二維流體,內部存在富含膽固醇、鞘脂和特定種類膜蛋白的微區,這一區域較厚且較少流動,被稱為“脂筏”;其如同一個蛋白質停泊的平台,與膜的信號轉導、蛋白質分選均有密切的關係
。
化學組成
質膜主要由膜脂和膜蛋白組成,另外還有少量糖,主要以糖脂和糖蛋白的形式存在。膜脂是膜的基本骨架,膜蛋白是膜功能的主要體現者。動物細胞膜通常含有等量的脂類和蛋白質。
質膜的流動性對生物體有很大影響。溫度、ph、脂分子結構、膽固醇等多方面影響流動性。古生菌的質膜是由單層或單雙層交替的磷脂分子組成,因此流動性較差,抗逆性強。
膜脂
膜脂主要包括磷脂、糖脂和膽固醇三種類型。
(一)、磷脂
是構成膜脂的基本成分,約占整個膜脂的50%以上。磷脂分子的主要特徵:
具有一個極性頭和兩個非極性的尾(脂肪酸鏈),但存在於線粒體內膜和某些細菌質膜上的心磷脂具有4個非極性的區域。
脂肪酸碳鏈為偶數,多數碳鏈由16,18或20個碳原子組成。常含有不飽和脂肪酸(如油酸)。
1、甘油磷脂
以甘油為骨架的磷脂類,在骨架上結合兩個脂肪酸鏈和一個磷酸基團,膽鹼、乙醇胺、絲氨酸或肌醇等分子籍磷酸基團連線到脂分子上。
主要類型有:磷脂醯膽鹼(phosphatidyl choline,PC,舊稱卵磷脂)、磷脂醯絲氨酸(phosphatidyl serine,PS)、磷脂醯乙醇胺(phosphatidyl ethanolamine ,PE,舊稱腦磷脂)磷脂醯肌醇(phosphatidyl inositol,PI)和雙磷脂醯甘油(DPG,舊稱心磷脂)等。
2、鞘磷脂
鞘磷脂(sphingomyelin,SM)在腦和神經細胞膜中特別豐富,亦稱神經醇磷脂,它是以鞘胺醇(sphingoine)為骨架,與一條脂肪酸鏈組成疏水尾部,親水頭部也含膽鹼與磷酸結合。原核細胞和植物中沒有鞘磷脂。
(二)、糖脂
糖脂(圖4-5、4-6)是含糖而不含磷酸的脂類,普遍存在於原核和真核細胞的質膜上,其含量約占膜脂總量的5%以下,在神經細胞膜上糖脂含量較高,約占5-10%。糖脂也是兩性分子。其結構與SM很相似,只是由一個或多個糖殘基代替了磷脂醯膽鹼而與鞘氨醇的羥基結合。
最簡單的糖脂是半乳糖腦苷脂,它只有一個半乳糖殘基作為極性頭部,在髓鞘的多層膜中含量豐富;變化最多、最複雜的糖脂是神經節苷脂,其頭部包含一個或幾個唾液酸和糖的殘基。神經節苷脂是神經元質膜中具有特徵性的成分。兒童所患的家族性白痴病(Tay-sachs disease)就是因為在其細胞內缺乏氨基己糖脂酶,不能將神經節苷脂GM2 加工成為GM3,結果大量的GM2累積在神經細胞中,導致中樞神經系統退化。神經節苷脂本身就是一類膜上的受體,已知破傷風毒素、霍亂毒素、干擾素、促甲狀腺素、絨毛膜促性腺激素和5-羥色胺等的受體就是不同的神經節苷脂。
(三)、膽固醇
膽固醇僅存在真核細胞膜上,含量一般不超過膜脂的1/3,植物細胞膜中含量較少,其功能是提高脂雙層的力學穩定性,調節脂雙層流動性,降低水溶性物質的通透性。如:在缺少膽固醇培養基中,不能合成膽固醇的突變細胞株很快發生自溶。
(四)、脂質體
質體(liposome)是一種人工膜。在水中磷脂分子親水頭部插入水中,疏水尾部伸向空氣,攪動後形成雙層脂分子的球形脂質體,直徑25~1000nm不等。脂質體可用於轉基因,或製備的藥物,利用脂質體可以和細胞膜融合的特點,將藥物送入細胞內部。
膜蛋白
膜蛋白是膜功能的主要體現者。據估計核基因組編碼的蛋白質中30%左右的為膜蛋白。根據膜蛋白與脂分子的結合方式,可分為整合蛋白(integral protein)、外周蛋白(peripheral protein)和脂錨定蛋白(lipid-anchored protein)。
整合蛋白可能全為跨膜蛋白(tansmembrane proteins),為兩性分子,疏水部分位於脂雙層內部,親水部分位於脂雙層外部。由於存在疏水結構域,整合蛋白與膜的結合非常緊密,只有用去垢劑(detergent)才能從膜上洗滌下來,如離子型去垢劑SDS,非離子型去垢劑Triton-X100。
蛋白的跨膜結構域可以是1至多個疏水的α螺鏇,形成親水通道的整合蛋白跨膜區域有兩種組成形式,一是由多個兩性α螺鏇組成親水通道;二是由兩性β摺疊組成親水通道。
外周蛋白靠離子鍵或其它較弱的鍵與膜表面的蛋白質分子或脂分子的親水部分結合,因此只要改變溶液的離子強度甚至提高溫度就可以從膜上分離下來,有時很難區分整合蛋白和外周蛋白,主要是因為一個蛋白質可以由多個亞基構成,有的亞基為跨膜蛋白,有的則結合在膜的外部。
脂錨定蛋白(lipid-anchored protein)可以分為兩類,一類是糖磷脂醯肌醇(glycophosphatidylinositol,GPI)連線的蛋白,GPI位於細胞膜的外小葉,用磷脂酶C(能識別含肌醇的磷脂)處理細胞,能釋放出結合的蛋白。許多細胞表面的受體、酶、細胞粘附分子和引起羊瘙癢病的PrPC都是這類蛋白。另一類脂錨定蛋白與插入質膜內小葉的長碳氫鏈結合,如三聚體GTP結合調節蛋白(trimeric GTP-binding regulatory protein)的α 和γ亞基。
特化結構
質膜常帶有許多特化的附屬結構。如:微絨毛、褶皺、纖毛、鞭毛等等,這些特化結構在細胞執行特定功能方面具有重要作用。由於其結構細微,多數只能在電鏡下觀察到。
(一)微絨毛
微絨毛(microvilli)是細胞表面伸出的細長指狀突起,廣泛存在於動物細胞表面。微絨毛直徑約為0.1μm。長度則因細胞種類和生理狀況不同而有所不同。小腸上皮細胞刷狀緣中的微絨毛,長度約為0.6~0.8μm。微絨毛的內芯由肌動蛋白絲束組成,肌動蛋白絲之間由許多微絨毛蛋白(villin)和絲束蛋白(fimbrin)組成的橫橋相連。微絨毛側面質膜有側臂與肌動蛋白絲束相連,從而將肌動蛋白絲束固定。
微絨毛的存在擴大了細胞的表面積,有利於細胞同外環境的物質交換。如小腸上的微絨毛,使細胞的表面積擴大了30倍,大大有利於大量吸收營養物質。不論微絨毛的長度還是數量,都與細胞的代謝強度有著相應的關係。例如腫瘤細胞,對葡萄糖和胺基酸的需求量都很大,因而大都帶有大量的微絨毛。
(二)皺褶
在細胞表面還有一種扁形突起,稱為皺褶(ruffle)或片足(lamllipodia)。皺褶在形態上不同於微絨毛,它寬而扁,寬度不等,厚度與微絨毛直徑相等,約0.1μm,高達幾微米。在巨噬細胞的表面上,普遍存在著皺褶結構,與吞噬顆粒物質有關。
(三)內褶
內褶(infolding,)是質膜由細胞表面內陷形成的結構,同樣具有擴大了細胞表面積的作用。這種結構常見於液體和離子交換活動比較旺盛的細胞。
(四)纖毛和鞭毛
纖毛(cilia)和鞭毛(flagella)是細胞表面伸出的條狀運動裝置。二者在發生和結構上並沒有什麼差別,均由9+2微管構成。有的細胞靠纖毛(如草履蟲)或鞭毛(如精子和眼蟲)在液體中穿行;有的細胞,如動物的某些上皮細胞,雖具有纖毛,但細胞本體不動,纖毛的擺動可推動物質越過細胞表面,進行物質運送,如氣管和輸卵管上皮細胞的表面纖毛。纖毛和鞭毛都來源於中心粒。關於纖毛和鞭毛的詳細結構和功能可參見第八章細胞骨架。
流動性
質膜的流動性由膜脂和蛋白質的分子運動兩個方面組成。
1.膜脂分子的運動
1. 側向擴散:同一平面上相鄰的脂分子交換位置(圖4-15)。
2. 鏇轉運動:膜脂分子圍繞與膜平面垂直的軸進行快速鏇轉。
3. 擺動運動:膜脂分子圍繞與膜平面垂直的軸進行左右擺動。
4. 伸縮震盪:脂肪酸鏈沿著與縱軸進行伸縮震盪運動。
5. 翻轉運動:膜脂分子從脂雙層的一層翻轉到另一層。是在翻轉酶(flippase)的催化下完成。
6. 鏇轉異構:脂肪酸鏈圍繞C-C鍵鏇轉,導致異構化運動。
2.影響膜流動性的因素
影響膜流動的因素主要來自膜本身的組分,遺傳因子及環境因子等。包括:
1. 膽固醇:膽固醇的含量增加會降低膜的流動性。
2. 脂肪酸鏈的飽和度:脂肪酸鏈所含雙鍵越多越不飽和,使膜流動性增加。
3. 脂肪酸鏈的鏈長:長鏈脂肪酸相變溫度高,膜流動性降低。
4. 卵磷脂/鞘磷脂:該比例高則膜流動性增加,是因為鞘磷脂粘度高於卵磷脂。
5. 其他因素:膜蛋白和膜脂的結合方式、溫度、酸鹼度、離子強度等。
3.膜蛋白的分子運動
主要有側向擴散和鏇轉擴散兩種運動方式。可用光脫色恢復技術(fluorescence recovery after photobleaching,FRAP)和細胞融合技術檢測側向擴散。鏇轉擴散指膜蛋白圍繞與膜平面垂直的軸進行鏇轉運動,膜蛋白的側向運動受細胞骨架的限制,破壞微絲的藥物如細胞鬆弛素B能促進膜蛋白的側向運動。
(1970年Larry Frye等人將人和和鼠的細胞膜用不同螢光抗體標記後,讓兩種細胞融合,雜種細胞一半發紅色螢光、另一半發綠色螢光,放置一段時間後發現兩種螢光抗體均勻分布。)
4.膜流動性的生理意義
質膜的流動性是保證其正常功能的必要條件。例如跨膜物質運輸、細胞信息傳遞、細胞識別、細胞免疫、細胞分化以及激素的作用等等都與膜的流動性密切相關。當膜的流動性低於一定的閾值時,許多酶的活動和跨膜運輸將停止,反之如果流動性過高,又會造成膜的溶解。
