
概述
中文名細胞譜系外文名celllineage命名者E·B·威爾遜命名時間1882年
這種細胞間在發育中世代相承的親緣關係猶如人類家族的譜系,故稱為細胞譜系。細胞譜系的研究對於了解卵質不均等分布和裂球發育命運的關係,以及比較不同種類動物早期發育之間的演化關係,都有重要作用。
簡史 1878年C.O.懷特曼研究螞蟥胚胎髮育時首先提出卵的卵裂是有序的過程,發育早期的每一裂球在構成身體時具有固定的形態學意義。1882年E.B.威爾遜創用了細胞譜系這一名詞。A.彭納斯在1922年對顫蚓胚胎的細胞譜系作了詳細的描述。
各種動物卵子的結構對其卵裂方式有直接影響。在不同卵裂方式下,細胞譜系中的命名方法也不相同。
輻射對稱型卵裂的細胞譜系
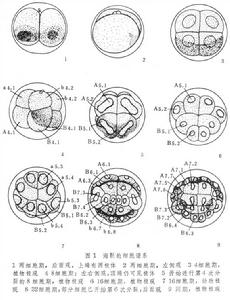 細胞譜系
細胞譜系以柄海鞘為例。E.G.康克林於1905年發現該動物受精卵植物性半球有含黃色物質的新月區即黃新月可作為標誌。第 1次卵裂面將黃新月區分為兩半,正好符合胚胎的對稱平面將卵分成相等的兩個裂球(AB2和AB2)。第2次卵裂面與第1次分裂面相垂直,將裂球增至4個,其中兩個在前、兩個在後,這就確定了胚胎的頭尾軸。前面的裂球稍大,名為A3和A3,後面的稍小,稱為B3和B3,黃色物質即在B3,B3中。第3次卵裂為緯裂,分成動物極4個小裂球和植物極四個大裂球,前者以小寫字母命名(a4.1,a4.2,b4.1和b4.2);後者以大寫字母命名(A4.1,A4.2,B4.1和B4.2)。第4次卵裂時動物極的小裂球為經裂,形成8個柱狀細胞;植物極的大裂球仍為緯裂,形成上下兩排,每排 4個細胞(圖1),黃色物質所在的4個大裂球此時居於胚胎後端。第5次卵裂為經裂,動物極的裂球都按前後分割;植物極的大裂球基本上按左右分割。由此可知海鞘卵的分裂,從一開始就體現了胚胎的對稱平面和頭尾方向。
螺旋型卵裂細胞譜系
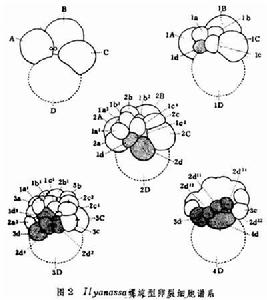 細胞譜系
細胞譜系特殊型式的卵裂細胞譜系
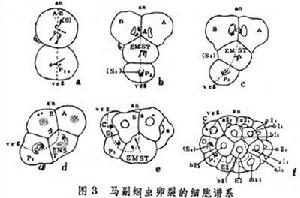 細胞譜系
細胞譜系在馬副蛔蟲卵裂中,絕大多數分裂球在生成過程中都要發生染色體消減,獨有成為原始生殖細胞的那個分裂球無此消減,這一點被引為種質連續學說的例證之一。
從60年代末期以來,一些分子生物學家十分注意發育和遺傳關係問題,因而對細胞譜系研究重新發生很大興趣。1973年德國分子生物學家 G.von埃倫施泰因用電鏡詳細地研究了一種自由生活的秀麗隱桿線蟲的細胞譜系。螞蟥的胚胎髮育,尤其是神經系統發育的細胞譜系也重新受到很大的重視。
B細胞譜系
哺乳動物的B細胞是在骨髓中發育成熟的,其發育階段經歷了祖B細胞(pro−Bcell)、大前B細胞(largepre−Bcell)、小前B細胞(smallpre−Bcell)、未成熟B細胞(immatureBcell)和成熟B細胞(matureBcell)或稱初始B細胞(naiveBcell)等幾個階段。B細胞發育過程中重要的生化事件幾乎都是圍繞著功能性BCR的表達和中樞免疫耐受的形成(圖12−1)。
①Ig重鏈、輕鏈基因重排:祖B細胞重鏈可變區基因先後發生D−J和V−DJ重排;到大前B細胞階段由於VDJ重排的完成,可表達完整的μ鏈,並作為pre−B細胞受體的一部分表達於大前B細胞表面;分化到小前B細胞階段,輕鏈的V−J發生重排;未成熟B細胞可表達完整的mIgM,只表達mIgM的未成熟B細胞部分可輸出骨髓,進入外周,如受到抗原刺激,則發生免疫耐受,這是B細胞中樞免疫耐受的主要機制;成熟B細胞同時表達mIgM和mIgD,抗原刺激後一般發生免疫正應答,使B細胞活化增殖,進一步分化為分泌Ig的漿細胞。
②Igα/Igβ:Igα(CD79a)和Igβ(CD79b)是BCR複合物的組成部分,主要介導抗原刺激後的信號傳遞。祖B細胞即開始表達Igα/Igβ異源二聚體,是B細胞重要標記。
③λ5和VpreB:大前B細胞表達pre−B受體,是由μ鏈和λ5/VpreB替代輕鏈所組成,是B細胞發育中一個重要的關卡點(checkpoint)。
