
物種形成
一個種在時間向度上的延續構成一個線系,種形成代表線繫上發生一個分枝;換句話說,一個分枝事件代表一個新種形成。但在某些情況下新種形成過程中並不發生分枝,即一個老種經過或長或短的進化過程就在形態上發生了顯著的改變,從而可以視為不同於老種的另一新種,這樣的種形成是線系進化的結果,所形成的種在古生物學上叫做“時間種”。種形成的方式 現代生物學關於種形成的研究對象大都是行有性生殖的動植物,因而種形成的研究多集中於“生殖隔離”的起源問題上,即研究在一個種的群體內如何產生或分化出與原群體生殖上隔離的亞群體,後者就是一個新種的開始。
C.R.達爾文早就注意到地理因素在種分歧和種形成中的作用。P.M.哈蒙德1981年將與種形成有關的地理因素區分為13種情況,並歸納為4種模式(見表)。
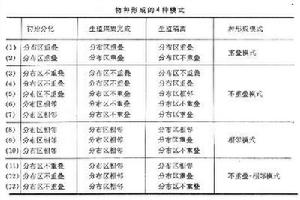 物種形成
物種形成此外,根據種形成過程的特點還可區分出兩種不同的種形成方式:①漸變的種形成,②量子種形成。
分布區不重疊和相鄰的種形成過程一般是漸變的,分布區重疊的種形成過程往往是“跳躍”的,即“量子種形成”方式。
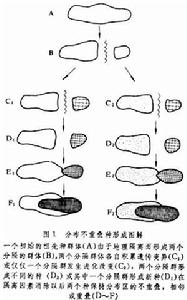
分布區不重疊的種形成 一個廣布的種,由於地理隔離因素而分隔為兩個或若干個相互隔離的群體,由於這些被隔離的群體之間的基因交流大大減弱或甚至完全停止,從而使各隔離群體之間的遺傳差異逐漸增加,通過若干中間階段(如形成族、亞種等)而最後達到生殖隔離。這樣,原先隔離的群體就形成不同的種。由於在初始分化和生殖隔離完成階段各個原群體的分布區不重疊, 故名不重疊種形成。 一旦生殖隔離完成,新種分布區即使再重疊 (例如由於地理隔離因素消失), 也不能交配或交配不育了,暗綠柳鶯便是一個很好的例子。此鳥分布很廣, 可分五個亞種, 形成為一個分布圈:①新疆亞種(P.t.viridanus),分布在中國新疆西部和北部, 國外伸展到中亞部分地區、西伯利亞西部直到歐洲北部; ②拉達亞種(P.t.ludlowi),分布在克什米爾地區,其北端與新疆亞種銜接,兩者相互雜交;③指名亞種(P.t.trochiloides),分布在中國西藏南部喜馬拉雅山地區,雲南西部和四川南部。其西端與拉達亞種銜接雜交;④甘青亞種(P.t.obscuratus),分布在中國甘肅、青海和四川西部,在四川和指名亞種相遇雜交;⑤東北亞種(P.t.plumbeitarsis),分布於中國東北、內蒙古和西伯利亞東部,其南端與甘青亞種間未發現有銜接區,其西部與新疆亞種在薩彥嶺一帶相遇,則不相雜交,已形成生殖隔離(圖1)。
 物種形成
物種形成在海洋生物中也有許多由於地理隔離而形成不同亞種的實例。某些物種在北太平洋東西兩側有不連續的分布,例如某些軟體動物和魚類在北美西海岸是一個亞種而在蘇聯遠東地區和日本北部沿海是同種的另一個亞種,原因是從白令海峽流過的來自北極的寒冷海流,構成了海洋生物的分布障礙。某些海洋動物在北大西洋和北太平洋對稱地分布而在北極中斷。例如鱈魚分布於北大西洋,在北太平洋則有同種的另一亞種,但在北極卻沒有鱈魚。顯然北極的寒冷是分布中斷的原因,但可能在某一地質時期北極地區曾經是比較溫暖的,兩大洋北部的生物曾有連續的分布。中美的某些生物,由於巴拿馬地峽的阻隔,而沿著東西兩岸分布著兩兩對稱的姐妹種。所謂姐妹種就是形態上非常相似而生殖上隔離的完全種。在地峽形成以前它們的分布可能是連續的。
分布區相鄰的種形成 分布於漸變的環境或不均一的環境中的廣布種,在分布區的不同部分形成相互接觸而又有一定程度的隔離(不完全的隔離)的群體。群體間在接觸區域有一定程度的基因交流,但群體的分布區的中心部分之間的基因交流很弱或甚至完全沒有。這樣,群體之間的遺傳差異會逐漸增大,由此而形成不同的種。某些間接的證據可以說明相鄰的種形成方式的存在。例如由呈環鏈狀分布的若干亞種組成的所謂“環布種”,相鄰的亞種之間有部分的基因交流,環鏈兩端的亞種之間可能分布重疊,但往往不能互相雜交或雜交不育(生殖隔離)。一個典型的實例就是環繞北極的鷗,它是由若干亞種組成的,更新世以後分布於北太平洋的亞種越過北美和北大西洋而與西歐的亞種相遇,但仍保持顯著的差異而不婚配 (圖2)。
 物種形成
物種形成分布區重疊的種形成 一些證據表明,生殖隔離可以由於生物種的行為差異、發情或開花季節差異、寄主,以及其他原因而導致在那些分布區並非完全不重疊的群體內產生。有些學者提出一些證據證明植物的某些新種的形成是沒有地理隔離的情況下達到生殖隔離的。
但分布區是否重疊往往很難區分。例如人體寄生有3種虱:陰虱寄生於腋毛和陰毛中,人虱分布於軀體(主要在衣服上),頭虱生於頭髮上。若從整個人體看,3種虱的分布區是重疊的,但若從人體的局部部位看,它們又是不重疊的。分布區是否重疊一般應根據具體的生物對象的活動能力和行為等特點來判斷。
分布區重疊的和相鄰的種形成方式 新種產生於遺傳上連續的單個群體之內(分布區重疊的種形成)或者產生於有基因交流的相鄰分布的一系列群體之中(分布區相鄰的種形成)。在沒有任何阻止基因交流的外在的隔離因素的情況下,新種從母群體中分化產生出來的一些證據多是從觀察研究“成對種”和“初始種”的分布和區分特徵而獲得的。“成對種”和“初始種”被認為是新形成的或形成中的物種,它們常是研究物種形成的對象。但是根據它們的地理分布和區分特徵來推斷種形成方式還存在許多問題。例如“成對種”或“初始種”的現有的分布區相鄰的狀況也可能是種間競爭所造成的地理分布上的相互排斥,如是,則其相鄰的分布乃是種形成的結果或種形成後的現象。母種與新種之間的區分特徵中哪些是種形成過程所涉及的,哪些是種形成以後產生的,也很難判斷。
根據學者們提供的資料,可將分布區重疊的和相鄰的種形成歸納為兩種可能的方式:①兩階段的種形成過程。在一個遺傳上連續的群體之內首先發生種內分異,經過相當長時間的種內分異的積累而最終在群體內產生生殖隔離的新種。②一步“跳躍”的過程。新種從母群體中突然產生,即通過染色體突變或基因突變或其它機制在較短時間內產生與母群基因交流阻斷的新類型或新種,一舉而達到生殖隔離。
關於第一種方式可以舉出下面一些事實。分布區重疊的種形成的實例如某些寄主專一的昆蟲寄主族是近期形成的半種,是種內分異的結果。
G.L.布希(1975)在研究果蠅的寄主族(指同一物種的有不同的寄主專一性的族群)後指出:有同一物種內寄主的轉換可以造成分布區重疊的寄主族。布希認為,對於果蠅來說寄主轉換隻需要很少的基因突變,而寄主的改變只是種內分異的第一步,其後是生態小生境的改變並伴隨寄主族之間、新寄主族與母群體之間的生殖的障礙。例如,果蠅的交配只發生在其寄主的果實上,不同的寄主族之間發生交配的可能性很小,這樣,寄主族之間的分異就會和地理隔離造成的效果一樣。
對澳洲蝗蟲的研究提供了分布區相鄰的種形成方式的例證。M.J.D.懷特(1978)研究了澳洲蝗蟲的相鄰分布的近緣種和染色體族之後提出了“原地靜態”的種形成方式,他認為,染色體重排(染色體融合、分離、倒位、易位、斷裂、缺失等)在種內初始分異中起重要作用。澳洲蝗蟲是失去飛翔能力的、種核心型變異很大的昆蟲,大約有 200種之多。在一些種 (例如在Vandiemenella viatica)之內有許多核型不同的、地理上相鄰分布的染色體族。有一些核型不同的近緣種的分布區也是相鄰的。這些相鄰分布的族或種的分布區往往有明確的邊界,互不重疊。研究還發現染色體族之間存在著程度不同的生殖障礙。例如,各族之間的雜合子不育或生育力低,從而在各染色體族之間形成一狹窄的“雜交帶”。由於基因交流的阻隔,染色體族有可能發展為生殖隔離的新種。
但對於這些分布區相鄰的染色體族和種的形成的解釋還有一些不同的看法。首先是關於染色體重排在種形成過程中究竟起多大作用還難以評價。在澳洲蝗蟲中並非所有的近緣種在核型上都有區別。另一個疑問是:相鄰的分布是否可能為種間競爭排斥的結果,即種形成後的現象。還有一個問題是:種內分異所造成的族(半種)之間的生殖障礙(不完全的生殖隔離)如何發展為完全的生殖隔離。達爾文和A.R.華萊士都曾提出過生殖障礙可以通過自然選擇而逐步發展的假說。如果種內分異產生出多種表型,如果中間類型(雜種)的適應值相對較低,那么通過“中間類型”或“雜種”的淘汰絕滅而由生殖障礙發展為完全的生殖隔離。
第二種跳躍的種形成方式是不經過種內分異而一步達到生殖隔離的過程。例如在植物中,多倍體的種可能是從其二倍體的親種中快速產生的。H.劉易斯(1966)認為木本植物的染色體基數在科或亞科級的大分類單元中是恆定的,在草本植物中染色體基數在屬一級的類群中是恆定的。植物的染色體基數的改變可能涉及高級分類群的形成,因而是跳躍的。
量子種形成 某些生物學家(例如G.G.辛普森,E.邁爾等)認為,除了漸變的種形成方式以外,還可能存在快速的、跳躍式的種形成方式。另一些生物學家(如斯坦利)則認為種形成不可能是漸變的過程,只能是“跳躍”式的、“相對快速”的過程。處於“相對穩定”狀態的種,通過“突然”的改變而分化出新種的過程稱為量子形成。這個詞最先由V.格蘭特1963年提出,S.M.斯坦利把它定義為“大部分進化集中發生在種形成的初期的相當短的時間(相對於種的壽命而言)之內的種形成過程”。關於種形成的方式是漸變論者與間斷平衡論者之間爭論的焦點。漸變論者認為漸變的種形成是一般的、普通的,快速、跳躍式的種形成是極特殊的情況;間斷平衡論者則認為種的形成只能是“跳躍”式的。
20世紀30年代,群體遺傳學家S.賴特1932年所提出的“適應峰”概念就包含有量子種形成的思想。賴特用類似地形圖的“適應圖”(圖3)來形象地表示生物種的基因組合對環境的適應狀態。賴特認為自然界中的種各自占據著適應圖中的“峰”(標有“+”號的部位),這些“峰”代表生物種的基因組合及其表型特徵具有最大適應值,而處於“谷”(標有“-”號的部位)中的基因組合適應值最低。由“峰”到“谷”之間的等高線代表適應值由高到低逐步下降。新種的形成意味著從一個 “峰”越過 “谷”而達到另一個“峰”,這隻有通過顯著的進化改變或快速的“跳躍”才能實現,因為緩慢的過渡要越過適應“谷”(適應值最低的狀態)是不可能的。
 物種形成
物種形成目前,關於量子種形成問題的爭論焦點在於快速種形成的具體機制。一些學者強調這種機制是“大突變”,甚至認為一個或少數幾個突變就能產生一個新種或新的高級分類單元。這實際上又回到了20世紀初期德·弗里斯的“突變論”。
有的學者則提出另一種快速種形成的可能機制:在分隔狀態下的小群體,隨機因素有助於快速進化和種形成。如果一個廣布的種由許多小群體組成,小群體之間有不完全的隔離,則隨機因素會造成小群體對總體的大的偏離和小群體之間的遺傳差異。但由於小群體的不穩定也增大了絕滅的可能性。
