探索歷程
發展原因
由於科學家在對物質跨膜運輸現象的研究中產生了一系列的疑問,於是開始從生物膜的功能深入探索到生物膜的結構。
脂質雙分子層
1895年,歐文頓(E. Overton)曾經用500多種化學物質對植物細胞的通透性進行過上萬次的實驗,發現細胞膜對不同物質的通透性不一樣:凡是可以溶於脂質的物質,比不能溶於脂質的物質更容易通過細胞膜進入細胞。於是他提出:膜是由脂質組成的。
20世紀初,科學家第一次將細胞膜從哺乳動物紅細胞中分離出來。化學分析表明,膜的主要成分是磷脂和蛋白質。1925年,荷蘭科學家戈特(E.Gorter)和格倫德爾(F.Grendel)用丙酮抽提紅細胞膜結構,計算出紅細胞膜平鋪面積同其表面積之比為(1.8~2.2)∶1,約為兩倍。他們由此提出脂質雙分子層模型。
單位膜模型
1935年,J. Danielli & H. Davson發現質膜的表面張力比油-水界面的張力低得多,推測膜中含有蛋白質,從而提出了“蛋白質-脂類-蛋白質”的三明治模型。認為質膜由雙層脂類分子及其內外表面附著的蛋白質構成的。1959年在上述基礎上提出了修正模型,認為膜上還具有貫穿脂雙層的蛋白質通道,供親水物質通過。
1959年,羅伯特森(J. D. Robertson)用超薄切片技術獲得了清晰的細胞膜照片,在電子顯微鏡下看到了細胞膜的暗-明-暗三層結構,厚約7.5nm,它由厚約3.5nm的雙層脂分子和內外表面各厚約2nm的蛋白質構成。他提出“單位膜模型”假說:連續的脂質雙分子層組成膜的主體,磷脂的非極性端朝向膜內側,極性端朝向膜外兩側,蛋白質以單層肽鏈的厚度,通過靜電作用與磷脂極性端相結合,從而形成蛋白質—磷脂—蛋白質的三層結構,稱之為單位膜。他提出真核細胞與原核細胞具有相同的膜結構。單位膜模型的主要不足在於:把生物膜的結構描述成靜止的、不變的,這顯然與膜功能的多樣性相矛盾。
流動鑲嵌模型
1970年,Larry Frye和Michael Lipids等科學家將人和鼠的細胞膜用不同螢光抗體標記後,讓兩種細胞融合,雜種細胞一半發紅色螢光、另一半發綠色螢光,放置一段時間後發現兩種螢光抗體均勻分布。這一實驗,以及相關的其他實驗證據表明細胞膜具有流動性。
在新的觀察和實驗證據的基礎上,又有學者提出了一些關於生物膜的分子結構模型。其中,1972年桑格(S. J. Singer)和尼克森(G. Nicolson)提出的流動鑲嵌模型
晶格鑲嵌模型
1975年,Wallach提出了晶格鑲嵌模型。他在流動鑲嵌模型的基礎上,進一步強調:生物膜中流動性脂質的可逆性變化。這種變化區域呈點狀分布在膜上。相變表現為膜脂分子的一種協同效益,即幾十個以上的脂分子同時相變。膜脂的相變受溫度、脂本身的性質、膜中其他成分、pH和二價陽離子濃度等因素的影響。
板塊鑲嵌模型
1977年,Jain和White提出了板塊鑲嵌模型,其內容本質上與晶格鑲嵌模型相同。他們認為:在流動的脂雙分子層中,存在許多大小不同的、剛度較大的、彼此獨立運動的脂質“板塊”(有序結構區),板塊之間被無序的流動的脂質區所分割,這兩種區域處於一種連續的動態平衡之中。
總結
生物膜的結構雖然有很多種,但被廣泛接受的結構模型基本內容是趨向一致的。其要點和特點基本相同,主要包括膜的分子組成和結構特徵。
主要內容
模型概述
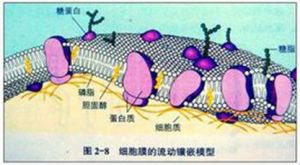
1972年,桑格(S. J. Singer)和尼克森(G. Nicolson)總結了當時有關膜結構模型及各種研究新技術的成就,提出了生物膜的流動鑲嵌模型(fluid mosaic model)。
流動鑲嵌模型(fluid mosaic model),特指膜的流動鑲嵌模型。這一模型是針對細胞膜提出的一種膜的結構模型,描述膜為結構和功能上不對稱的脂雙層所組成,蛋白質以鑲嵌樣模式分布在膜的表面與內部,並能在膜內運動。此模型也可適用於亞細胞結構的膜。
基本理論
生物膜的流動鑲嵌模型(fluid mosaic model)認為:
一、磷脂雙分子層構成了生物膜的基本支架,這個支架不是靜止的。其中磷脂分子的親水性頭部朝向兩側,疏水親脂性的尾部相對朝向內側。
二、球形膜蛋白分子以各種鑲嵌形式與磷脂雙分子層相結合,有的鑲在磷脂雙分子層表面,有的全部或部分嵌入磷脂雙分子層中,有的貫穿於整個磷脂雙分子層。這裡體現了膜結構內外的不對稱性。另外,大多數膜蛋白分子是功能蛋白。
三、大多數蛋白質分子和磷脂分子都能夠以進行橫向擴散的形式運動,體現了膜具有一定的流動性。
四、在細胞膜的外表,有一層由細胞膜上的蛋白質與糖類結合形成的糖蛋白,叫做糖被。它在細胞生命活動中具有重要的功能。例如:消化道和呼吸道上皮細胞表面的糖蛋白有保護和潤滑作用;糖被與細胞表面的識別有密切的關係,好比是細胞與細胞之間,或者細胞與其他大分子之間,互相聯絡用的文字或語言。除糖蛋白外,細胞膜表面還有糖類和脂質分子結合成的糖脂。
主要特點
流動鑲嵌模型有兩個主要特點。第一個特點是,蛋白質不是伸展的片層,而是以摺疊的球形鑲嵌在磷脂雙分子層中,蛋白質與膜脂的結合程度取決於膜蛋白中胺基酸的性質。第二個特點是,膜具有一定的流動性,不再是封閉的片狀結構,以適應細胞各種功能的需要。
綜合評價
這一模型強調了膜結構的流動性和不對稱性,對細胞膜的結構和功能作出了較為科學的解釋,被廣泛接受,也得到許多實驗的支持。
流動鑲嵌模型在某些方面還不夠完善,如忽略了無機離子和水所起的作用,忽視了蛋白質分子對膜脂分子流動性的控制作用,忽視了膜的各個部分流動性的不均勻性等等。
結構特性
鑲嵌性
磷脂雙分子層和蛋白質的鑲嵌面;或按二維排成相互交替的鑲嵌面。
流動性
流動性概述
膜的流動性是細胞膜結構的基本特徵之一,同時也是細胞膜表現其正常功能的必要條件。膜的流動性是指膜結構分子的運動性,它包括膜脂的運動和膜蛋白的運動。多年來,人們藉助於不斷創新的實驗技術,對於膜結構特性的研究進一步加深,並不斷取得進展。
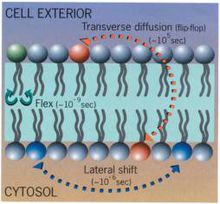 膜脂的運動
膜脂的運動膜脂的流動性
在正常生理狀況下,膜脂分子處於運動狀態。膜脂的運動方式主要有側向擴散、鏇轉運動、鏇轉異構運動、左右擺動以及翻轉運動等。膜脂的流動性受著一些因素的影響,主要影響因素有:
①溫度:在一定溫度下,脂分子從液晶態(能流動具有一定形狀和體積的物態)轉變為凝膠狀(不流動)的晶態。 這一能引起物相變化的溫度稱為相變溫度。 當環境溫度在相變溫度以上時,膜脂分子處於流動的液晶態; 而在相變溫度以下時, 則處於不流動的晶態。 膜脂相變溫度越低,膜脂流動性就越大;反之,相變溫度越高,膜脂的流動性也就越小。
②膜脂的脂肪酸鏈:飽和程度高的脂肪酸鏈因緊密有序地排列,因而流動性小;而不飽和脂肪酸鏈由於不飽和鍵的存在,使分子間排列疏鬆而無序,相變溫度降低,從而增強了膜的流動性。脂肪酸鏈的長度對膜脂的流動性也有影響:隨著脂肪酸鏈的增長,鏈尾相互作用的機會增多,易於凝集(相變溫度增高),流動性下降。
③膽固醇:膽固醇對膜脂流動性的調節作用隨溫度的不同而改變。在相變溫度以上,它能使磷脂的脂肪酸鏈的運動性減弱,從而降低膜脂的流動性。而在相變溫度以下時,膽固醇可通過阻止磷脂脂肪酸鏈的相互作用,緩解低溫所引起的膜脂流動性劇烈下降。
除以上因素外,膜脂與膜蛋白的結合程度、環境中的離子強度、pH 值等都會影響膜脂的流動性。
膜脂的流動是造成細胞膜流動的主要因素,概括起來,膜脂的運動方式主要有六種。
① 側向擴散(lateral diffusion);
② 鏇轉運動(rotation);
③ 伸縮運動(flex);
④ 翻轉擴散(transverse diffusion), 又稱為翻轉(flip-flop);
⑤ 左右擺動;
⑥ 鏇轉異構運動。
膜蛋白的流動性
細胞膜中的蛋白質也能以側向擴散等方式運動。人們通過實驗已充分證實了膜蛋白的流動性。David Frye和 Michael Edidin利用螢光抗體免疫標記來測定細胞表面抗原的分布。 首先用綠色螢光標記的抗體處理培養的小鼠細胞, 用紅色螢光標記的另一種抗體處理人細胞。 由於抗體與細胞膜上抗原的特異性結合, 使得處理後的小鼠細胞和人細胞分別發出綠色、紅色螢光。在滅活的仙台病毒介導作用下,小鼠細胞與人細胞融合,所形成的雜交細胞一半呈綠色一半呈紅色。在 37℃溫育40 分鐘後,雙色螢光均勻分布於雜交細胞表面。螢光點的均勻分布,主要是由於結合有螢光抗體的抗原在膜平面經側向擴散而重新分布。
膜蛋白的運動也受很多因素影響。 膜中蛋白質與脂類的相互作用、 內在蛋白與外在蛋白相互作用、膜蛋白複合體的形成、膜蛋白與細胞骨架的作用等都影響和限制蛋白質的流動。
膜蛋白主要有以下幾種運動形式:
① 隨機移動 有些蛋白質能夠在整個膜上隨機移動。移動的速率比用人工脂雙層測得的要低。
② 定向移動 有些蛋白比較特別,在膜中作定向移動。例如,有些膜蛋白在膜上可以從細胞的頭部移向尾部。
③ 局部擴散 有些蛋白雖然能夠在膜上自由擴散,但只能在局部範圍內擴散。
不對稱性
概述
細胞質膜的不對稱性(membrane asymmetry)是指細胞質膜脂雙層中各種成分不是均勻分布的,包括種類和數量的不均勻。(以脂雙層分子的疏水端為界,生物膜可分為近胞質面和非胞質面內外兩層。生物膜內外二層的結構和功能有很大差異,這種差異稱為生物膜的不對稱性。)
膜的主要成分是蛋白、脂和糖,膜的不對稱性主要是指這些成分分布的不對稱以及這些分子在方向上的不對稱。
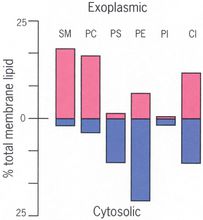 膜脂的不對稱性
膜脂的不對稱性膜脂的不對稱性
膜脂的不對稱性表現在脂雙層中分布的各類脂的比例不同,各種細胞的膜脂不對稱性差異很大。
膜蛋白的不對稱
每種膜蛋白在膜中都有特定的排布方向,與其功能相適應,這是膜蛋白不對稱性的主要因素。膜蛋白的不對稱性包括外周蛋白分布的不對稱以及整合蛋白內外兩側胺基酸殘基數目的不對稱。
 膜糖分布的不對稱性
膜糖分布的不對稱性膜糖的不對稱性
膜糖以糖蛋白或糖脂的形式存在,無論是糖蛋白還是糖脂的糖基都是位於膜的外表面。
不對稱性的意義
膜脂、膜蛋白及膜糖分布的不對稱性導致了膜功能的不對稱性和方向性,保證了生命活動的高度有序性。
