基因盒為附著在一個小的識別部位的一個耐藥基因組成,數個基因盒可以被包裝成一個多基因盒陣列,並依次被整合進入一個易於快速流動的較大的DNA單位,稱為整合子。
結構
整合子是一種遺傳因素,包含一個能捕獲外源基因的位點特異重組系統,這些被捕獲的外源基因通常是耐藥基因(如編碼氨基糖苷類、β-內醯胺類、氯黴素、磺胺類等的基因)。這些耐藥基因被包含在叫做基因盒的單一的移動單元。整合子由兩個保守片段即5′端和3′端保守段組成,在5′端保守段包含有編碼整合酶的Int基因、Int特異重組位點(attI)和負責基因轉錄的啟動子P1(Pant)和P2,但P2 僅少見於少數整合子。Int位於整合子5′保守末端,屬於酪氨酸整合酶家族,其編碼的整合酶識別兩個不同的重組整合位點,即位於整合子上的attⅠ位點和位於基因盒的59-be家族成員(attc位點)。整合酶負責催化attⅠ位點和一個59-be位點之間、兩個59-be位點之間及兩個attⅠ位點之間的重組,但兩個attⅠ位點之間的重組效率比attⅠ位點和一個59-be位點之間的重組效率低。3′端保守段的結構則因整合子類型的不同而異。基因盒是單一的可移動的DNA分子,通常以環行獨立的狀態存在。只有當它被整合子捕獲並整合到整合子中才能轉錄。基因盒通常不含啟動子,但一旦基因盒插入整合子,這個基因就能在5′端的共同啟動子Pant作用下轉錄。基因盒是由一個ORF(通常是耐藥基因)和一個反向不完全重複序列即59-be組成,59-be片段交換位點長度從50 bp ~150 bp不等。在反向重複序列的5′端有一個長7 bp反向核心位點RYYYAAC;在3′端有一個核心位點GTTRRRY(R為嘌呤,Y為嘧啶)。基因盒插入attⅠ位點導致基因盒下游的此級位點(attc)的形成。一個整合子可以捕獲一個或多個基因盒,被捕獲的基因盒5′端與attⅠ位點,3′端的59-be片段與attc位點發生特異重組。整合子-基因盒結構如圖1。
分類
目前已發現和鑑定的整合子有10種類型, 但只有5種是和編碼抗生素耐藥性的基因盒相關, 其 中Ⅰ類 ~Ⅲ類研究的較多,而且在耐藥性傳遞方面起著重要作用。
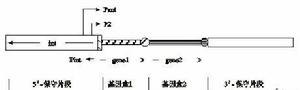 圖1整合子結構圖
圖1整合子結構圖Ⅰ類整合子是最常見的,其結構包括5′和3′CS和一個內部可變區(VR)編碼抗生素耐藥基因。對大多數Ⅰ型整合子,5′保守段均相似,包括intI和attI,然而3′- CS存在不同,包括3個開放閱讀框(ORF);季銨鹽化合物解放溴乙錠的耐藥基因(quacE△1),磺胺耐藥基因和一個未知功能的ORF5。
Ⅱ類整合子位於轉座子Tn7及其衍生物上,其基因序列與Ⅰ類整合子非常相似,是缺陷的intI基因,其編碼的整合酶與一類整合酶有46%的同源性。目前Ⅱ類整合子上的耐藥基因有核苷轉移酶基因aadA1a,甲氧胺嘧啶基因核鏈絲黴素基因(sat)。
Ⅲ類整合子是在耐亞胺培南的黏質沙雷菌的耐藥質粒上發現的,其整合酶編碼320個胺基酸與整合酶Ⅰ有51%同源性。最近一種新型的Ⅲ類整合子被發現。這類整合子長2 863 bp,由intI 3整合酶基因、attI3重組位點、兩個啟動子合兩個基因盒組成IntI3整合酶合黏質沙雷菌AK 9 373整合子種的整合酶有98.8%同源性。Bla-GES-1基因盒插入attI1位點,第二個基因盒是bla-OXA-10和aac(6′)Ib基因盒融合的結果,編碼對卡那黴素的耐藥性。
Ⅳ類整合子是一類特殊的整合子,他是由Didier Mazel 等1997年從霍亂弧菌基因組中檢測出來的,全長126 kb,包含至少179個基因,被稱為超整合子。最近超整合子在不同的細菌種中被發現。超整合子基因編碼蛋白的單一的適應功能,包括耐藥性、毒力、新陳代謝活動等很好地解釋了其為宿主適應性方面提供了很多好處。在微生物適應性方面的影響最驚人的體現是它快速廣泛地出現在相似抗生素耐藥性的描述。
移動
整合子攜帶著重組的基因盒插入到轉座子或接合質粒中,在不同的細菌中運動而傳播耐藥性。另外,整合子能捕獲和切除基因盒,一個整合子可以捕獲一個和多個基因盒,且同一個基因盒可以整合到不同的整合子上,從而導致基因盒的移動。
整合酶識別兩個重組位點:一個是存在於整合子上的attI位點,另一個是存在於基因盒上59-be家族(attc)位點。attI位點和59-be位點不同,59-be家族包含了許多不同長度不同序列的的位點,而不同類型的整合子的attI位點具有相同的序列,attI1、attI2、attI3被檢測到僅僅只有一個潛在的單一位點。在59-be和attI位點發生重組時可形成3種不同形式的反應:attI×59-be, 59-be×59-be,attI×attI,這3種形式的反應都被IntI催化,但attI×attI反應的頻率很低。 有關基因盒的整合以Ⅰ型整合子為例,如圖2所示,游離的基因盒以閉環狀結構存在,通過整合酶IntI1催化以線形結構結合於attI1和attc位點,其整合過程是可逆的。在整合線形基因被切下後,由於兩端的特殊結構又回復到環狀,並可整合到其他整合子上。並且多個基因盒可以整合到整合子中,多重插入導致整合子多重基因盒排列的形成。在染色體整合子中可以插入150多個基因盒。
表達
JP2包含克隆整合子片段的質粒是不同的,其差異在於啟動子的不同以及整合子基因盒系統中表達耐藥性的基因盒的數量和順序的不同。大多數基因盒不含啟動子,檢測所有轉錄產物發現有共同的啟動子Pant。所有基因盒都順著其編碼區一個方向插入,從而保證它們以共同的啟動子Pant表達。Pant是惟一的與大腸桿菌啟動子具有很強相似性的區域。Pant序列的改變影響基因盒所表達的耐藥性的水平。當Pant和P2 同時存在時,兩個啟動子的轉錄產物都可被檢測到;當不止一個基因盒存在時,則基因盒的位置影響其表達的耐藥性的水平,當基因位於第一個基因盒,靠近Pant時,耐藥性表達水平最高。隨著基因盒距離的增加,其表達水平逐漸下降。
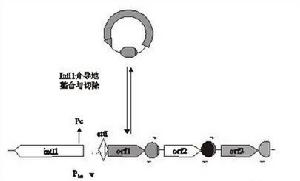 圖2基因盒的整合
圖2基因盒的整合抗生素耐藥基因表達水平受許多因素影響,其中最明顯的是轉錄和翻譯的起始信號。耐藥基因通常位於質粒上,同樣的基因也可以在不同的質粒中發現,因此質粒的拷貝數也是一個因素。有研究表明,具有35-TTGACA序列和10TAAACT序列的Pant,其表達活性要低20倍。在一些自然的整合子中發現單獨存在的Pant的弱形式,但在In2(Tn21),Pant和下游的次級啟動子P2一起發揮作用,從而補充弱Pant的低活性,它們一起表達的鏈黴素抗性基因僅比強Pant低3倍。
