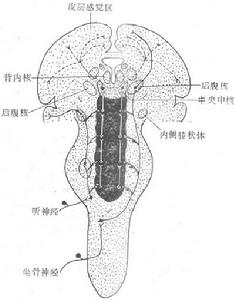
 感覺投射系統
感覺投射系統概述
一般認為,經典的感覺傳導首是由三個神經元的接替完成的。第一級神經元位於脊髓神經節或有關的腦神經感覺神經節內,第二級神經元位於脊髓后角或腦幹的有關神經核內,第三級神經元就在丘腦的感覺接替核內。但特殊感覺(視、聽、嗅)的傳導道情況比較複雜。視覺傳導道包括視桿及視錐細胞在內,則為四個神經元接替;聽覺傳導道更為複雜,從外周到大腦皮層很難說包含幾個神經元接替;嗅覺傳導道與丘腦感覺接替核無關。所以,一般經典的感覺傳導道就是通過丘腦的特異投射系統而後作用於大腦皮層的;它們都投身到大腦皮層的特定區域,每一種感覺的傳導投射系統都是專一的,各種感覺上傳都有其專門的途徑。
圖10-23 感覺投射系統示意圖
黑色區代表腦幹網狀結構 實線代表丘腦
特異投射系統 虛線代表丘腦非特異投射系統
自從對腦幹網狀結構的研究開展以來,逐步認識到感覺傳導向大腦皮層投射還有另一條途徑。那就是當上述經典傳導道的第二級神經元纖維通過腦幹時,發出其側支與腦幹網狀結構內神經元發生突觸聯繫;然後在網狀結構內反覆換元上行,抵達丘腦的第三類細胞群,進一步向大腦皮層作彌散性投射。所以,這一感覺投射途徑就是通過丘腦的非特異投射系統而後作用於大腦皮層的。這一投射系統是不同感覺的共同上傳途徑,也就是說當不同感覺傳入腦幹部分由側支進入網狀結構後,就不再是專一特異的傳導系統,而是由同一上行系統向上傳導。電生理研究支持這一結論,當記錄腦幹網狀結構內單個神經元電活動時,來自不同部位的感覺刺激傳入衝動都可激海參或抑制同一細胞的電活動,說明在網狀中感覺投射具有聚合的性質。
特異投射系統年行纖維進入大腦皮層的第四層後,形成絲球結構主要與第四層內神經元形成突觸聯繫,通過若干中間神經元接替,轉而與大錐體細胞的細胞體形成突觸聯繫,誘發其興奮。非特異性投射系統的上行纖維進入皮層後分散在各層,以自由末端的方式與皮層神經元的樹突形成突觸聯繫,改變其興奮狀態。以微電極研究大腦皮層神經元單位放電的實驗,可以證明這一點。例如,當刺激丘腦後腹核時,大腦皮層感覺區內的神經元通常只放電一次,即使增加刺激強度也能使神經元持續放電超過二三次;當刺激丘腦第三類細胞群時,則很難使大腦皮層感覺區神經放電,但台在這刺激之後緊接著刺激丘腦後腹核,神經元便會連續放電四五次以上。由此可見,非特異投射系統本身不能單獨激發皮層神經元放電,但可改變大腦皮層的興奮狀態。
非特異投射系統的功能,還可在另一些實驗中觀察。例如,刺激動物中腦網狀結構,能喚醒動物,腦電波呈去同步化快波;而在中腦頭端中斷網狀結構時,出現類似睡眠的現象,腦電波呈現同步化慢波(圖10-24)。由此說明,在腦幹網狀結構內具有上行喚醒作用的功能系統,這一系統稱為腦幹網狀結構上行激動系統(ascending reticular activaring system)。目前知道,上行激動系統主要就是通過丘腦非特異投射系統而發揮作用的,其作用就是維持與改變大腦皮層的興奮狀態。由於這一系統是一個多突觸接替的上行系統,因此易於受藥物的影響而發生傳導阻滯。例如,巴比妥類催眠藥作用可能就是由於阻斷了上行激動系統的傳導;一些全身麻醉藥(如乙醚)也可能是首先抑制了上行激動系統和大腦皮層的活動而發揮麻醉作用的。
圖10-24切斷特異性傳導道或非特異性傳導道後貓的行為與腦電圖變化
A為切斷特異性傳導道而不損傷非特異性傳導道的貓,處於覺醒狀態A′為其腦電圖
B為切斷非特異性傳導道的貓,處於昏睡狀態 B′為其腦電圖
