
後生動物
正文
動物界除原生動物門以外的所有多細胞動物門類的總稱。包括多孔動物門、腔腸動物門、扁形動物門等(見無脊椎動物)。其特徵是體軀由大量形態有分化、機能有分工的細胞構成;與群體原生動物的兼有營養和生殖功能的細胞不同,其生殖細胞和營養細胞有明顯的分化。依體制形態的對稱情況,後生動物可分為不對稱動物(多孔動物門)、輻射對稱動物(腔腸動物門、櫛水母動物門、棘皮動物門;後者的對稱是次生的、櫛水母和某些珊瑚是左右輻射對稱)和兩側對稱動物(其他所有門類)。後生動物在胚胎髮育過程中有胚層的分化,其中多孔動物門只有內胚層和外胚層的初步分化,腔腸動物門在內外胚層間又有中膠層。自扁形動物門以後的門類都是三胚層動物。
根據體腔的有無和結構(圖1)
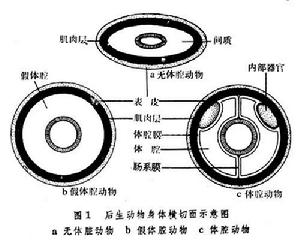 可將後生動物分為無體腔動物,包括多孔動物門、腔腸動物門和扁形動物門;假體腔動物,包括線形動物門、腹毛動物門等(見無脊椎動物);體腔動物(Coelomata),包括環節動物以後的所有動物門類。
可將後生動物分為無體腔動物,包括多孔動物門、腔腸動物門和扁形動物門;假體腔動物,包括線形動物門、腹毛動物門等(見無脊椎動物);體腔動物(Coelomata),包括環節動物以後的所有動物門類。 後生動物根據形態結構可分為4個相應的水平:①實囊胚級水平,如中生動物有一些只是由一層細胞以及內部的空腔組成,另一些是由比較少數的細胞擁擠在一起形成實心的、管狀的體軀或者形成一個由許多細胞以及若干層細胞所組成的板狀構造。由於這些動物的細胞排列與高等動物的囊胚期的排列相似,故有人稱之為實囊幼蟲型動物。②細胞水平,如多孔動物門由二層細胞,即外面的皮層和裡面的胃層構成。身體的各種機能由或多或少獨立生活的細胞如領細胞完成。③組織級水平,在組織內不僅有細胞,也有非細胞形態的物質(基質、纖維等)。如腔腸動物開始分化出上皮組織(具有神經一樣的傳導功能)等。④器官系統級水平,從扁形動物門起,動物有了不同細胞,不同組織組成的結構和機能不同的器官系統。
在後生動物中,紐蟲最先具備完整的消化管,一端為口,另一端為肛門。此外,消化管與循環系統分離,神經系統也比扁形動物更高等。
古生物學、形態學和胚胎學的證據表明:多細胞的後生動物來自單細胞的原生動物。在多細胞動物尚未興起時,單細胞動物已盛極一時。單細胞動物群體與多細胞動物之間並沒有絕對的鴻溝,由許多小的細胞(有時細胞數可達50000個)組成的空球狀的團藻蟲就是介於單細胞和多細胞之間的中間類型。
單細胞動物通過什麼樣的方式進化為多細胞動物,各派意見不一。主要學說有:
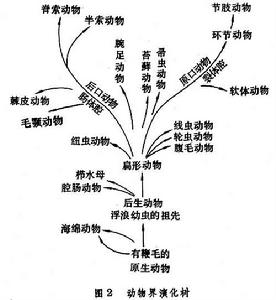
①原腸蟲學說,1874年E.海克爾首先提出多細胞動物最早的祖先是類似團藻的球形群體,這些單細胞動物在某一面內陷形成多細胞動物的祖先。因為它們和原腸胚很相似,有兩胚層和原口,所以海克爾稱之為原腸蟲,把他的學說稱之為原腸蟲學說。②吞噬蟲學說,N.N.梅契尼科夫觀察很多低等細胞動物的胚胎髮育,發現了一些較低等的種類,其原腸主要不是由內陷方法,而是由內移方法形成的。他還發現某些多細胞動物主要是靠吞噬作用進行細胞內消化,很少為細胞外消化。因此,他推論最初出現的多細胞動物是進行細胞內消化,細胞外消化是後來才發展的。在類似團藻蟲這樣的單細胞群體中,某些細胞吞噬食物後進入群體之內形成內胚層,結果形成二胚層動物,起初為實心,後來才逐漸地形成消化腔。因此,梅契尼柯夫便把這種假想的多細胞動物的祖先叫做吞噬蟲。③合胞體學說,J.哈奇和E.D.漢森認為後生動物的祖先應是和多核纖毛蟲相似的一種單細胞動物,這些多核纖毛蟲的細胞膜分割演變為多細胞動物。這個學說認為扁形動物中無腸類是最原始的後生動物。由原始的多核纖毛蟲一支進化為兩側對稱的、有纖毛的、但無大小核之分的無腸類,另一支則演變成有大小核之分的高等纖毛蟲(圖2)。
 後生動物
後生動物江靜波等:《無脊椎動物學》,人民教育出版社,北京,1982。
