
基本功能
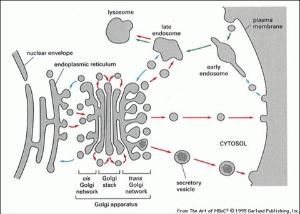 細胞的生物膜系統
細胞的生物膜系統1.分室作用細胞的膜系統不僅把細胞與外界環境隔開,而且把細胞內的空間分隔,使細胞內部的區域化(compartmentation),即形成各種細胞器,從而使細胞的代謝活動“按室進行”。各區域內均具特定的pH、電位、離子強度和酶系等。
同時,由於內膜系統的存在,又將各個細胞器聯繫起來共同完成各種連續的生理生化反應,比如光呼吸過程就是由葉綠體、過氧化物體和線粒體三者協同完成的。
2.代謝反應的場所細胞內的許多生理生化過程在膜上有序進行。如光合作用的光能吸收、電子傳遞和光合磷酸化、呼吸作用的電子傳遞及氧化磷酸化過程分別是在葉綠體的光合膜和線粒體內膜上進行的。
3.物質交換質膜的另一個重要特性是對物質的透過具有選擇性,控制膜內外進行物質交換。如質膜可通過擴散、離子通道、主動運輸及內吞外排等方式來控制物質進出細胞。各種細胞器上的膜也通過類似方式控制其小區域與胞質進行物質交換。
4.識別功能質膜上的多糖鏈分布於其外表面,似“觸角”一樣能夠識別外界物質,並可接受外界的某種刺激或信號,使細胞作出相應的反應。例如,花粉粒外壁的糖蛋白與柱頭細胞質膜的蛋白質之間就可進行識別反應。膜上還存在著各種各樣的受體(receptor),能感應刺激,傳導信息,調控代謝。
化學組成
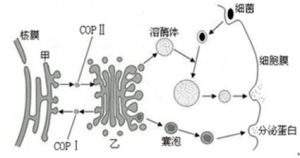 細胞的生物膜系統
細胞的生物膜系統這一比例關係反映到生物膜結構上,就是脂類以雙分子層構成生物膜的基本結構,而蛋白質分子則“鑲嵌”於其中。
(一)膜蛋白
生物膜中的蛋白質約占細胞蛋白總量的20%~30%,它們或是單純的蛋白質,或是與糖、脂結合形成的結合蛋白。根據它們與膜脂相互作用的方式及其在膜中的排列部位,可以大體地將膜蛋白分為兩類:外在蛋白與內在蛋白-。外在蛋白(extrinsicprotein)為水溶性球狀蛋白質,通過靜電作用及離子鍵等非共價鍵與膜脂相連,分布在膜的內外表面。內在蛋白(intrinsicprotein)占膜蛋白總量的70%~80%,又叫嵌入蛋白或整合蛋白,其主要特徵是水不溶性,分布在脂質雙分子層中,有的橫跨全膜也稱跨膜蛋白(transmembraneprotein),有的全部埋入疏水區,有的與外在蛋白結合以多酶複合體形式與膜脂結合。最近,又在生物中發現一類新的膜蛋白,叫膜脂蛋白,它們的蛋白部分不直接嵌入膜,而依賴所含的脂肪酸插入脂質雙分子層中。
膜蛋白執行著生物膜的主要功能。不同生物膜所具有的不同生物學功能主要是由於所含膜蛋白的種類和數量的不同。
(二)膜脂
在植物細胞中,構成生物膜的脂類主要是複合脂類(complexlipids),包括磷脂、糖脂、硫脂等。
磷脂(phospholipid)是含磷酸基的複合脂。在植物細胞膜中重要的磷脂屬甘油磷脂,它們是磷脂醯膽鹼(卵磷脂,phosphatidylcholine)和磷脂醯乙醇胺(腦磷脂,phosphatidylethanolamine)。另外,還有磷脂醯絲氨酸(phosphatidylserine),磷脂醯甘油(phosphatidylglycerinele),磷脂醯肌醇(phosphatidylinositol)等。
磷脂分子結構既有疏水基團,又有親水基團。
如圖所示,分子中有一個極性的“頭部”和一個疏水的“尾部”。磷脂的這種特性使之在生物膜形成中起著獨特的作用。 糖脂(glycolipid)是指甘油脂中甘油分子上有一個羥基以糖苷鍵與一分子六碳糖相結合的產物。硫脂(sulpholipid)則是糖脂分子中的六碳糖上又帶一個硫酸根基團。糖脂和硫脂也具有極性的“頭部”和疏水的“尾部”,這兩種脂類在葉綠體膜中特別多,其含量甚至超過了磷脂。
由上可知,膜上的脂類幾乎都是兩性分子,在水相中可自發地形成脂雙層,即脂類分子呈兩層排列,親水的頭部處於水相,疏水的尾部朝向中央。這種自發的排列過程稱作脂類的自我裝配。脂雙層一旦有破損也能自我閉合。實驗表明:脂雙層是流動的,脂類分子能在各自的單分子層內迅速地移動,即橫向擴散,而一般不容易“翻轉”,即不易從一個單分子層顛轉到另一單分子層。脂雙層的自我裝配、自我閉合以及具有流動性的三大特點決定了它能成為生物膜理想的基本結構。
膜脂上的脂肪酸(fattyacid)有飽和脂肪酸和不飽和脂肪酸之分,不飽和脂肪酸分子有雙鍵,其順式和反式的互變使不飽和脂肪酸易於彎曲或轉動,從而使得膜結構比較鬆散而不僵硬。
膜脂上的不飽和脂肪酸與植物的抗逆性有很大關係,通常耐寒性強的植物,其膜脂中不飽和脂肪酸含量較高,而且不飽和程度(雙鍵數目)也較高,有利於保持膜在低溫時的流動性;而抗熱性強的植物,其飽和脂肪酸的含量較高,有利於保持膜在高溫時的穩定性。
(三)膜糖
生物膜中的糖類主要分布於質膜的外單分子層。這些糖是不超過15個單糖殘基所連線成的具分支的低聚糖鏈(寡糖鏈),它們大多數與膜蛋白共價結合,少部分與膜脂結合,分別形成糖蛋白和糖脂(圖1-7)。由於單糖彼此間結合方式、排列順序、種類、數量以及有無分支等差別,其組合是千變萬化的,所形成的寡糖鏈種類非常多,形成了多種細胞表面特異的圖像,細胞之間藉此進行互相識別和交換信息。
主要結構
關於生物膜的結構有許多假說與模型,下面介紹兩種模型。
(一)流動鑲嵌模型
流動鑲嵌模型(fluidmosaicmodel)由辛格爾(S.J.Singer)和尼柯爾森(G.Nicolson)在1972年提出,認為液態的脂質雙分子層中鑲嵌著可移動的蛋白質,圖1-7展示了此模型的結構特點。內在蛋白嵌合在磷脂分子層中,內在蛋白或其聚合體可橫穿膜層,兩端極性部分伸向水相,中間疏水部分與脂肪酸部分呈疏水結合,外在蛋白與膜兩側的極性部分結合。
這個模型的特點是強調膜的不對稱性和流動性。不對稱性主要是由脂類和蛋白質分布的不對稱造成的。雖然同一種磷脂可見於脂雙層的任一層,但它們的數量是不等的。蛋白質在膜中有的半埋於內分子層,有的半埋於外分子層,即使貫穿全膜的蛋白質也是不對稱的。另外,寡糖鏈的分布也是不對稱的,它們大多分布於外分子層。膜的流動性包含兩個方面,其一是脂類分子是液晶態可動的,脂類分子隨溫度改變經常處於液晶態和液態的動態平衡之中,兩相中脂類分子排列不同,流動性大小也不同。其二是分布於膜脂雙分子層的蛋白質也是流動的,它們可以在脂分子層中側向擴散,但不能翻轉擴散。這說明了少量膜脂與膜蛋白有相對專一的作用,這種作用是膜蛋白行使功能所必須的。
脂雙層的流動性保證了膜能經受一定程度的形變而不致破裂,這可使膜中各種成分按需要調整或組合,使之合理分布,有利於表現膜的多種功能。更重要的是它允許膜互相融合而不失去膜對通透性的控制,確保膜分子在細胞分裂時均等地分配給子代細胞。如果膜不具有流動性,則很難想像細胞如何存活、生長和繁殖。流動鑲嵌模型雖得到比較廣泛的支持,但仍有很多局限性,如忽視了蛋白質對脂類分子流動性的控制作用和膜各部分流動的不均勻性等問題。
(二)板塊鑲嵌模型
板塊鑲嵌模型(platemosaicmodel)由賈因(M.K.Gain)和懷特(White)在1977年提出。他們認為,由於生物膜脂質可以在環境溫度或其它化學成分變化的影響下,或是由於膜中同時存在著不同脂質(脂肪鏈的長短或不同的飽和度),或者由於蛋白質和蛋白質、蛋白質和脂質間的相互作用,使膜脂的局部經常處於一種“相變”狀態,即一部分脂區表現為從液晶態轉變為晶態,而另一部分脂區表現為從晶態轉變為液晶態。因此,整個生物膜可以看成是由不同組織結構、不同大小、不同性質、不同流動性的可移動的“板塊”所組成,高度流動性的區域和流動性比較小的區域可以同時存在,隨著生理狀態和環境條件的改變,這些“板塊”之間可以彼此轉化。
