
物質概述
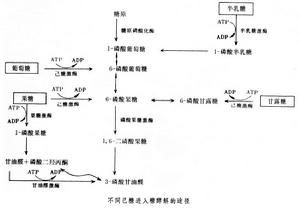 糖酵解
糖酵解丙酮酸的進一步代謝,因生物種屬的不同以及供氧情況的差別而有不同的道路。例如在無氧情況下,強烈收縮的動物肌肉細胞中,丙酮酸還原為乳酸,在許多微生物中可分解為乙醇或乙酸等;在有氧情況下,則氧化成二氧化碳和水。
具體過程
糖酵解可分為二個階段,活化階段和放能階段。準備階段
(1)葡萄糖磷酸化(Phosphorylation)“葡萄糖氧化”是放能反應,但“葡萄糖”是較穩定的化合物,要使之放能就必須給予“活化能”來推動此反應,即必須先使“葡萄糖”從“穩定狀態”變為“活躍狀態”,活化1個葡萄糖需要消耗1個ATP——由ATP放出1個高能磷酸鍵,約放出30.5KJ自由能,大部分變為熱量而散失,小部分使磷酸與葡萄糖結合生成“葡萄糖-6-磷酸”。催化酶為“己糖激酶”,此反應必須有Mg2+的存在。
(2)“葡萄糖-6-磷酸”重排生成“果糖-6-磷酸”。催化酶為“葡萄糖磷酸異構酶”。
(3)“果糖-6-磷酸”經酶催化生成“果糖-1,6-二磷酸"。催化酶為"1,6-磷酸果糖激酶"。
同(1)步驟一樣,此步反應再消耗1分子ATP。此步同樣是"ATP的γ-磷酸基團"經酶的作用轉移到底物上生成目標產物。
(4)“果糖-1,6-二磷酸”斷裂成“3-磷酸甘油醛”(glyceraldehyde 3-phosphate)和“磷酸二羥丙酮”,催化酶為“醛縮酶”。
(5)“磷酸二羥丙酮”很快被酶催化為“3-磷酸甘油醛”。催化酶為“丙糖磷酸異構酶”。
以上為第一階段,1個6C的葡萄糖轉化為2個3C化合物PGAL(phosphoglyceraldehyde),消耗2個ATP用於葡萄糖的活化。
另一種不是由葡萄糖為初始底物進入EMP的方式——以“葡萄糖-1-磷酸”形式進入EMP,則僅消耗1個ATP。
無論是以上哪種進入方式,在這一階段都沒有發生氧化還原反應。
放能階段
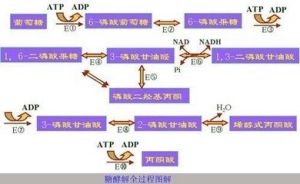
(6)“3-磷酸甘油醛”氧化生成“1,3-二磷酸甘油酸”(1,3-diphosphoglycerate),釋放出2個e-和1個H+,傳遞給電子受體NAD+,生成NADH和H+,並且將能量轉移到高能磷酸鍵中。催化酶為“3-磷酸甘油醛脫氫酶”。(7)不穩定的“1,3-二磷酸甘油酸”失去高能磷酸鍵,生成“3-磷酸甘油酸”(3-phosphoglycerate),能量轉移到ATP中,1個“1,3-二磷酸甘油酸”生成“1個ATP”。催化酶為“磷酸甘油酸激酶”。
【此步驟中發生第一次底物水平磷酸化】
(8)“3-磷酸甘油酸”重排生成“2-磷酸甘油酸”(2-phosphoglycerate)。催化酶為“磷酸甘油酸變位酶”。
(9)“2-磷酸甘油酸”脫水生成“磷酸烯醇式丙酮酸”——PEP(phospho-enol-pyruvate)。催化酶為“烯醇化酶”。
(10)PEP將磷酸基團轉移給ADP生成ATP,同時形成丙酮酸。催化酶為丙酮酸激酶。
【此步驟中發生第二次底物水平磷酸化】
以上為糖酵解第二個階段。1分子的PGAL在酶的作用下生成1分子的丙酮酸。在此過程中,發生一次氧化反應生成一個分子的NADH,發生兩次底物水平的磷酸化,生成2分子的ATP。這樣,一個葡萄糖分子在糖酵解的第二階段共生成4個ATP和2個NADH和H+,產物為2個丙酮酸。
在糖酵解的第一階段,1個葡萄糖分子活化中要消耗2個ATP。因此在糖酵解過程中1個葡萄糖生成2分子的丙酮酸的同時,淨得2分子ATP和2分子NADH和H+,NADH和H+通過不同的穿梭途徑進入到線粒體參與呼吸鏈,產生不同數量的ATP(α-磷酸甘油穿梭將H交給FAD,後產生1.5個ATP;蘋果酸-天冬氨酸穿梭將H+交給NADH+H+,後產生2.5個ATP)。
調節
正常生理條件下,人體內的各種代謝過程受到嚴格而精細的調節,以保持內環境穩定,適應機體生理活動的需要。這種調節控制主要是通過改變酶的活性來實現的。己糖激酶(葡萄糖激酶)、磷酸果糖激酶-1、丙酮酸激酶是糖酵解的關鍵酶,它們的活性大小,直接影響著整個代謝途徑的速度和方向,其中以磷酸果糖激酶-1最為重要。
1.激素的調節胰島素可誘導GK、PFK-1、PK的合成,因而使糖酵解過程增強。
2.代謝物對限速酶的變構調節磷酸果糖激酶-1(PFK-1)是三個限速酶中催化效率最低的,故而是糖酵解途徑中最重要的調節點。該酶分子為四聚體。分子中不僅具有與底物結合的部位,還具有與變構激活劑和變構抑制劑結合的部位。F-1,6-BP、ADP、AMP等是其變構激活劑,而ATP、檸檬酸等為其變構抑制劑。在這些代謝物的共同調節下,機體可根據能量需求調整糖分解速度。當細胞內能量消耗增多,ATP濃度降低,AMP、ADP濃度增加,則磷酸果糖激酶-1被激活,糖分解速度加快,使ATP生成量增加;當細胞內有足夠的ATP儲備時,ATP濃度增加,AMP、ADP濃度下降,磷酸果糖激酶-1被抑制,糖分解速度減慢,減少ATP生成量,避免能量的浪費;當飢餓時,機體動員儲存脂肪分解氧化,生成大量乙醯CoA,乙醯CoA可與草醯乙酸縮合成檸檬酸,抑制磷酸果糖激酶-1的活性,從而減少糖的分解,以維持飢餓狀態下血糖濃度。
操作步驟
糖酵解的第一步是葡萄糖磷酸化為6-磷酸葡萄糖。不同細胞類型中所含有的酶也不一樣,在所有的細胞中,皆有己糖激酶(Hexokinase)進行催化,而在肝細胞和胰腺中,則另外含有一種稱為葡(萄)糖激酶(Hexokinase IV)的酵素。磷酸化過程消耗一分子ATP,後面的過程證明,這是回報很豐厚的投資。細胞膜對葡萄糖通透,但對磷酸化產物6-磷酸葡萄糖不通透,後者在細胞內積聚並繼續反應,將反應平衡向有利於葡萄糖吸收的那一面推移。之後6-磷酸葡萄糖會在磷酸己糖異構酶的催化下生成6-磷酸果糖。(在此果糖也可通過磷酸化進入糖酵解途徑)接著6-磷酸果糖會在磷酸果糖激酶的作用下被一分子ATP磷酸化生成1,6-二磷酸果糖,ATP則變為ADP。這裡的能量消耗是值得的,:首先此步反應使得糖酵解不可逆地繼續進行下去,另外,兩個磷酸基團可以進一步在醛縮酶的參與下分解為磷酸二羥丙酮和3-磷酸甘油醛。磷酸二羥丙酮會在磷酸丙糖異構酶幫助下轉化為3-磷酸甘油醛。兩分子3-磷酸甘油醛會被NAD+和3-磷酸甘油醛脫氫酶(GAPDH)的氧化下生成1,3-二磷酸甘油酸(1,3-BPG)。
下一步反應,1,3-二磷酸甘油酸轉變為3-磷酸甘油酸。此反應由磷酸甘油酸激酶催化,高能磷酸鍵由1,3-二磷酸甘油酸轉移到ADP上,生成兩分子ATP。在此,糖酵解能量盈虧平衡。兩分子ATP消耗了又重新生成。ATP的合成需要ADP作原料。如果細胞內ATP多(ADP則會少),反應會在此步暫停,直到有足夠的ADP。這種反饋調節和重要,因為ATP就是不被使用,也會很快分解。反饋調節避免生產過量的ATP,節省了能量。磷酸甘油酸變位酶推動3-磷酸甘油酸生成2-磷酸甘油酸,最終成為磷酸烯醇式丙酮酸。磷酸烯醇式丙酮酸是高能化合物。最後,在丙酮酸激酶的作用下磷酸烯醇式丙酮酸生成一分子ATP和丙酮酸。此步反應也受ADP調節。
糖酵解中的不可逆反應
人體可通過糖異生,即從非糖化合物,如丙酮酸和乳酸等物質重新合成葡萄糖。當肝或腎以丙酮酸為原料進行糖異生時,糖異生中的其中七步反應是糖酵解中的逆反應,它們有相同的酶催化。但是糖酵解中有三步反應,是不可逆反應。在糖異生時必須繞過這三步反應,代價是更多的能量消耗。這三步反應都是強放熱反應,它們分別是:
1.葡萄糖經己糖激酶催化生成6磷酸葡萄糖ΔG=-33.5kJ/mol
2.6磷酸果糖經磷酸果糖激酶催化生成1,6二磷酸果糖ΔG=-22.2kJ/mol
3.磷酸烯醇式丙酮酸經丙酮酸激酶生成丙酮酸ΔG=-16.7kJ/mol
糖酵解中的調節位點
糖酵解在體內可被精確調節,這樣一方面可以滿足機體對能量的需要,另一方面又不會造成浪費。同時,當細胞內還進行糖異生的時候,調節就顯得非常重要了,因為要避免空循環的發生。調節是通過改變底物濃度,酶的活性實現的。
磷酸果糖激酶是其中最重要的限速酶,這也是巴斯德效應的關鍵參與者,它也決定了糖異生的速度,成為調節位點。AMP的濃度越高,酶的活性越高。就是當機體大量消耗了ATP,而相應又產生了很多AMP的時候,酶的活性提高,使得糖酵解按生成ATP的方向快速前進,以提高ATP產量。
生理意義
糖酵解可以把釋放的自由能轉移到ATP中。糖酵解也是果糖、甘露糖、半乳糖等己糖的共同降解途徑。果糖及甘露糖通過己糖激酶的催化作用可轉變成果糖-6-磷酸,果糖還可以通過一系列酶的作用轉變成3-磷酸甘油醛。半乳糖可以在一些酶催化下轉變成1-磷酸葡萄糖。有些先天性代謝疾病是由於上述果糖與半乳糖代謝中的某些酶缺失所致。如缺失磷酸果糖醛縮酶,則果糖-1-磷酸在肝、腸及腎中堆積引起肝腫大及肝腎及腸吸收功能衰退,患這種病的兒童不能服用果糖或蔗糖。
能量轉化
平衡點
值得一提的是,生成1,6-二磷酸果糖後的大部分反應都是向能量升高的方向進行的,沒有酶(磷酸果糖激酶(PFK),磷酸甘油酸激酶(PGK))的催化,是不會自發進行的。而糖酵解的逆過程--糖異生(從甘油等非糖物質生成葡萄糖)則容易進行,此過程用到大部分在糖酵解裡面出現過的酶,除了提到的兩位“車夫”外,它們只出現在糖酵解中。在糖異生這兩步逆反應會放出大量的熱,分別為-14及-24kJ/mol。無氧環境和有氧環境
在糖酵解中,每分子葡萄糖提供兩分子ATP。真核生物的線粒體能同時從兩分子丙酮酸中另外獲得36分子ATP。能量轉化的多少取決於在細胞質中產生的NADH+H通過線粒體膜的方式。不論在無氧還是有氧環境中,糖酵解成丙酮酸這一過程都能進行。3-磷酸甘油醛在3-磷酸甘油醛脫氫酶GAPDH的作用下脫氫。脫下的氫離子會將氧化劑(輔酶)NAD還原成NADH+H。NAD會在呼吸鏈中再生。若在無氧環境,放熱的(ΔG´=-25kJ/mol)乳糖脫氫酶(LDH)反應會再生NAD:丙酮酸的還原會生成乳糖和再生NAD(酵母則會使用另外兩種酶—丙酮酸脫羧酶加乙醇脫氫酶)。無氧環境下糖酵解GAPDH-和LDH-反應的相互聯繫,除了少部分NADH+H會被磷酸甘油脫氫酶(GDH)轉化外,大部分會用於再生NAD。
關鍵酶
糖酵解的關鍵酶:有3個,即己糖激酶、6-磷酸果糖激酶-1和丙酮酸激酶,它們催化的反應基本上都是不可逆的。重要性
6-磷酸果糖激酶-1>丙酮酸激酶>己糖激酶ATP/AMP比值的高低對6-磷酸果糖激酶-1活性的調節有重要意義。當ATP濃度較高時,6-磷酸果糖激酶-1幾乎無活性,糖酵解作用減弱;當AMP累積,ATP較少時,酶活性恢復,糖酵解作用加強;此外,H+也可抑制6-磷酸果糖激酶-1的活性,這樣可防止肌肉中形成過量的乳酸。
發現
1897年,德國生化學家E.畢希納發現離開活體的釀酶具有活性以後,極大地促進了生物體內糖代謝的研究。釀酶發現後的幾年之內,就揭示了糖酵解是動植物和微生物體內普遍存在的過程。英國的F.G.霍普金斯等於1907年發現肌肉收縮同乳酸生成有直接關係。英國生理學家A.V.希爾,德國的生物化學家O.邁爾霍夫、O.瓦爾堡等許多科學家經歷了約20年,從每一個具體的化學變化及其所需用的酶、輔酶以及化學能的傳遞等各方面進行探討,於1935年終於闡明了從葡萄糖(6碳)轉變其中乳酸(3碳)或酒精(2碳)經歷的12箇中間步驟,並且闡明在這過程中有幾種酶、輔酶和ATP等參加反應。意義
 糖酵解
糖酵解2.通過糖酵解使葡萄糖降解生成ATP,為生命活動提供部分能量,尤其對厭氧生物是獲得能量的主要方式
3.糖酵解途徑為其他代謝途徑提供中間產物(提供碳骨架),如6-磷酸葡萄糖是磷酸戊糖途徑的底物;磷酸二羥丙酮?a-磷酸甘油?合成脂肪
4.是糖有氧分解的準備階段
5.由非糖物質轉變為糖的異生途徑基本為之逆過程
