
基本信息
 發光動物
發光動物發光動物涉及的門類有:原生動物發光放射蟲;腔腸動物(線形美螅、仙遊水母、燐海鰓海 、燐瓜水母);紐形動物(燐紐蟲);環節動物(燐沙蠶、發光蚯蚓);軟體動物(殼斗海筍 Pholadidea ponita、發光海牛、橙海牛、發光蝸牛、螢烏賊 Watasentia scintillans、雙喙耳烏賊Sepiola briostrata Sasahi);
 螢火蟲
螢火蟲甲殼類(發光性橈足類10多種、赫氏海螢Cypridina hig-endorfi);多足類(發光蜈蚣);昆蟲類(螢科昆蟲數十種、螢叩頭蟲、雌光螢科的大場雌光螢、蕈蠅科的螢火蚋;螢火蟲幼蟲也會發光);外肛動物(棘膜苔蟲Acan-thodesia serrata);棘皮動物(發光性陽遂足6種);
原索動物(柱頭蟲Balanoglossidae、燐火體蟲Pyrosoma atlanticum、紐鰓樽Salpa);魚類(亮烏賊 Etmopteus lucifer、裸鰮Diaphus coeruleus、日本松球魚Monocentris japonica、提燈 Cory-nolophus reinhardii等200種以上)等(除上述外,還有蛙和守宮也有能自己發光的物質)。發光有由自身產生髮光物質而自己發光的一次發光,以及由共生者相互依賴的共生髮光或發光共生(德Leuchtsymbiose),即二次發光。這兩種發光是有區別的。
分類
 螢火蟲幼蟲
螢火蟲幼蟲具有生物發光能力的動物。在動物界的分布是分散而無系統,涉及的門類有:原生動物(夜光蟲、Pyrocystis、發光放射蟲);腔腸動物(線形美螅、仙遊水母、燐海鰓Pennaluta phosphorea、海仙人掌Cavernularia obesa Moroff、燐瓜水母);紐形動物(燐紐蟲);環節動物(燐沙蠶、發光蚯蚓);軟體動物(殼斗海筍 Pholadidea ponita、發光海牛、橙海牛、發光蝸牛、螢烏賊 Watasentia scintillans、雙喙耳烏賊Sepiola briostrata Sasahi);甲殼類(發光性橈足類10多
種、赫氏海螢Cypridina hig-endorfi);多足類(發光蜈蚣);昆蟲類(螢科昆蟲數十種、螢叩頭蟲);外肛動物(棘膜苔蟲Acan-thodesia serrata);棘皮動物(發光性陽遂足6種);原索動物(柱頭蟲Balanoglos
 發光動物
發光動物sidae、燐火體蟲Pyrosoma atlanticum、紐鰓樽Salpa);魚類(亮烏賊 Etmopteus lucifer、裸鰮Diaphus coeruleus、日本松球魚Monocentris japonica、提燈Cory-nolophus reinhardii等200種以上)等(除上述外,還有蛙和守宮也有能自己發光的物質)。發光的形式和發光裝置,因種類不同而異。發光有由自身產生髮光物質而自己發光的一次發光,以及由共生者相互依賴的共生髮光或發光共生(德Leuchtsymbiose),即二次發光。這兩種發光是有區別的。共生或寄生的發光,主要是由於發光細菌的發生和寄生,但也有因游沙蠶的附著而使Crateromorpha(海綿的一種)發光,以及由於燐紐蟲的寄生使柄海鞘發光的例子。許多寄生髮光的魚類的發光,是連續性的發光,而自身發光動物的發光,常常是間歇性的發光;發光細胞對外界刺激(夜光蟲、海仙人掌)和神經刺激(螢蟲、螢烏賊)的反應,可出現瞬間性的閃光,平時則維持低光度的光亮。發光的神經控制,對螢蟲等則是與雌雄求偶(信號刺激)的閃光作用有密切關係。發光在夜行性動物、深海動物、洞穴動物中是很多的,但發光的生物學意義卻不十定都很清楚,就發光烏賊來說,有雌雄引誘說、外敵防禦說、恐嚇說等不同的學說;深海魚類,除此以外,還有一種在頭上高高掛著可動性的“觸手”,其頂端的發光器能一明一暗地發光,提燈就說是這種例子,對此有兩種學說,即獵物誘惑和暗中照明。
共生或寄生的發光
 發光動物
發光動物主要 是由於發光細菌的發生和寄生,但也有因游沙蠶的附著而使Crateromorpha(海綿的一種)發光,以及由於燐紐蟲的寄生使柄海鞘發光的例子。許多寄生髮光的魚類的發光,是連續性的發光,而自身發光動物的發光,常常是間歇性的發光;發光細胞對外界刺激
 發光動物
發光動物(夜光蟲、海仙人掌)和神經刺激(螢蟲、螢烏賊)的反應,可出現瞬間性的閃光,平時則維持低光度的光亮。發光的神經控制,對螢蟲等則是與雌雄求偶(信號刺激)的閃光作用有密切關係
 發光動物
發光動物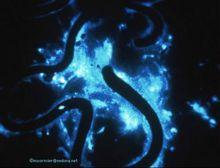 發光動物
發光動物作用
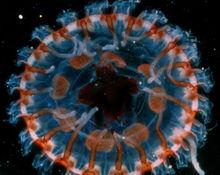 發光動物
發光動物發光在夜行性動物、深海動物、洞穴動物中是很多的,但發光的生物學意義卻不十定都很清楚,就發光烏賊來說,有雌雄引誘說、外敵防禦說、恐嚇說等不同的學說;深海魚類,除此以外,還有一種在頭上高高掛著可動性的“觸手”,其頂端的發光器能一明一暗地發光,提燈就說是這種例子,對此有兩種學說,即獵物誘惑和暗中 照明。
