
過程
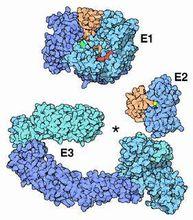 泛素化
泛素化具體過程:泛素化修飾涉及泛素激活酶 E1、泛素結合酶 E2和泛素連線酶 E3的一系列反應:首先在 ATP(紅色所示)供能的情況下酶 E1(蛋白質編號1r4n)粘附在泛素分子尾部(淡黃色所示)的 Cys殘基上(綠色所示,注意在這個結構中,Cys突變為Ala)激活泛素,接著,E1將激活的泛素分子轉移到E2酶上(蛋白質編號1fxt),隨後, E2酶和一些種類不同的E3酶共同識別靶蛋白,對其進行泛素化修飾。根據 E3與靶蛋白的相對比例可以將靶蛋白單泛素化修飾和多聚泛素化修飾。 E3酶(蛋白質編號1ldk和1fqv)的外形就像一個夾子,靶蛋白連線在中間的空隙內(星號所示)。酶的左側結構域決定靶蛋白的特異性識別,右側結構域定位E2酶以轉移泛素分子。蛋白質泛素化的結果是使得被標記的蛋白質被蛋白酶分解為較小的多肽、胺基酸以及可以重複使用的泛素。
蛋白質降解
泛素-蛋白酶體途徑是先發現的,也是較普遍的一種內源蛋白降解方式。需要降解的蛋白先被泛素化修飾,然後被蛋白酶體降解。
不過後來又發現,並非所有泛素化修飾都會導致降解。有些泛素化會改變蛋白的活性,導致其他的生物效應,如DNA損傷修復,機體免疫應答等。
蛋白質泛素化
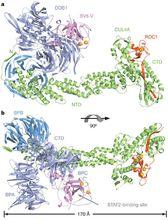 泛素蛋白
泛素蛋白蛋白質泛素化作用是後翻譯修飾的一種常見形式,該過程能夠調節不同細胞途徑中各式各樣的蛋白質底物。通過一個三酶級聯(E1-E2-E3),蛋白質的泛素連線由E3泛素連線酶催化,這種酶是cullin-RING複合體超級家族的最佳代表。
在從酵母到人類的各級生物中都保守的DDB1-CUL4-ROC1複合體是最近確定出的cullin-RING泛素連線酶,這種酶調節DNA的修復、DNA複製和轉錄,它能被病毒所破壞。
由於缺少一個規則的SKP1類cullin連線器和一種確定的底物召集結構域,目前人們還不清楚DDB1-CUL4-ROC1 E3複合體如何被裝配起來以對各種蛋白質底物進行泛素化。
在這項新的研究中,DDB1-CUL4A-ROC1複合體被病毒劫持的形式進行了晶體結構分析。分析結果表明DDB1利用一個β-propeller結構域作為cullin骨架結合物,利用一種多變的、附著的獨立雙β-propeller摺疊來進行底物的呈遞。
通過對人類的DDB1和CUL4A複合體進行聯繫提純,然後進行質譜分析,研究人員確定出了一種新穎的WD40-repeat蛋白家族,這類蛋白直接與DDB1的雙propeller摺疊結合併充當E3酶的底物募集模組。這些結構和蛋白質組學研究結果揭示出了cullin-RING E3複合體的一個新家族的裝配和多功能型背後的結構機制和分子邏輯關係。
泛素化類型
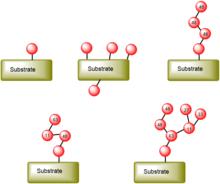 泛素化
泛素化E1,E2,E3對底物的泛素化可形成幾種不同的泛素化底物。有的底物蛋白只能被單泛素化,如 H2B;有的底物蛋白有多個賴氨酸殘基,在合適條件下會被多位點單泛素化;還有一些蛋白在單個賴氨酸位點會形成多聚泛素鏈,這種多聚泛素鏈可以根據連線泛素鏈的賴氨酸位點的不同可以分為單一、混合以及樹枝狀的結構。
作用
p53穩定性的變化與其功能調解密切相關 ,嚴密調節p53的代謝穩定性對正常細胞的生長發育非常重要。p53為一短半衰期轉錄因子 , p53蛋白的轉換由泛素依賴的蛋白水解途徑調節[3]。本研究結果表明 ,p53N端與降解有關的片段不能與其轉錄活性片段相分離 ;抑制或破壞泛素蛋白酶體水解通路對轉錄反應無普遍作用 ,但可特異性抑制p53的轉錄反應。本研究非常有意義的發現是 ,在蛋白酶體抑制劑存在或泛素化途徑缺失時 , p53穩定性增強 ,細胞內p53蛋白水平增加 ,然而p53的轉錄活性卻降低。即泛素蛋白酶體水解途徑被抑制時 , p53的轉錄功能也受到破壞 ,說明p53的蛋白水解途徑對其轉錄反應是必須的 ,兩過程存在功能性聯繫。此發現提示 ,泛素蛋白酶體調節的蛋白降解通路在p53轉錄中發揮重要的作用。以往研究中 ,在甲狀腺受體及雌激素調節的轉錄過程也發現同樣的規律[1 0 , 1 1 ]。我們推測 ,至少在某些細胞內活性蛋白 ,泛素蛋白酶體水解通路在轉錄激活及基因表達調節中發揮重要的作用。該發現對於細胞生長、分化的調節及疾病的診治 ,如保留p53的活性 ,預防和治療腫瘤將具有重要的意義。
p53泛素蛋白水解過程與轉錄激活過程間的生化學聯繫 ,還不甚清楚。研究支持以下的構想 ,某些細胞因子連線並參與這兩個過程。有研究表明轉錄輔助激活因子p3 0 0 /CBP在p53的轉錄反應中發揮重要的作用,而p3 0 0 /mdm2複合體還參與mdm2調節的p53降解反應 , p53、mdm2、p3 0 0C/H1特異性的相互作用為mdm2調節的p53轉換過程的重要步驟[1 3]。我們在近期研究中發現 ,p3 0 0通過連線泛素化和降解過程 ,在mdm2調節的p53轉換中發揮重要的作用[6]。上述結果提示 ,轉錄激活因子p3 0 0 /CBP在 p53泛素蛋白水解與轉錄激活過程的細胞內聯繫中 ,具有重大意義。當然我們也 不能排除其它細胞因子如mdm2等在該過程中所起的作用 ,詳盡的分子機制有待於進一步的研究。
----文章來自(解剖科學進展 2002年第4期第卷 大連醫科大學藥理教研室;美國俄亥俄州立大學細胞生物化學系;大連醫科大學生物化學教研室 遼寧大連116027 姚繼紅;Qianzheng Zhu;A A Wani;楊佩滿;崔秀雲)
Damaged DNA時,激活ATM,活化的ATM使p53發生磷酸化,而激活。磷酸化的p53使細胞在G1期出現生長停滯,進行DNA修復。如修復失敗,則通過活化bax基因使細胞進入凋亡,以保證基因組的遺傳穩定。
使依賴P53的周期素依賴激酶(CDK)抑制者P21和DNA修復基因(growth arrest and DNA damage 45,GASS45)上調性轉錄。註:GASS45圖中未標識出來。
Rb蛋白通過脫磷酸而活化,與E2F家族結合,阻斷了DNA上的S期基因的轉錄。反之,磷酸化的Rb蛋白與E2F分離,使得E2F與DP1蛋白形成異二聚體,活化S期基因的轉錄。註:DP1未標出。
Cyclin D1通過與CDK4 激酶作用使Rb 蛋白磷酸化,調節細胞周期G1~S 的過渡,是G1期細胞增殖的關鍵蛋白。PCNA促進此致癌的過程。
bcl-2(B-cell lymphoma/leukemia,bcl),這一家族的成員主要是抑制細胞凋亡而不是促進細胞增殖。
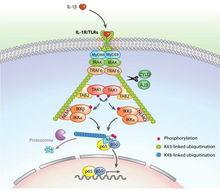 泛素化
泛素化