
簡介
肩帶的演化
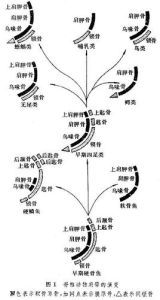 帶骨
帶骨四足動物的肩帶
現代四足動物的肩帶中,屬於軟骨原骨的基本上由3塊骨片組成,即肩胛骨、烏喙骨和前烏喙骨。3對軟骨原骨中,穩定少變的只有肩胛骨,其他兩對變化較大,有的兩者都缺;加入的膜原骨在原始種類數目多,以後逐步減少只剩下鎖骨,有些哺乳動物連鎖骨也沒有。
有尾兩棲動物的肩帶
有尾兩棲動物的肩帶大部分仍為軟骨,僅在肩臼附近開始骨化,完全缺少膜原骨,鎖骨也不存在。無尾兩棲動物(蛙或蟾蜍)的肩帶由肩臼分為背、腹兩部。背部為肩胛骨及上肩胛骨,腹部靠後面的是烏喙骨,由烏喙骨之內側向前延伸出上烏喙骨。蟾蜍兩側的上烏喙骨彼此交叉,叫弧胸型;蛙的上烏喙骨不交叉,相互平行癒合在一起,叫固胸型。肩帶的類型屬弧胸型還是固胸型是無尾兩棲動物分類標準之一。上烏喙骨之前為鎖骨,將前烏喙骨包在其中。在鎖骨、肩胛骨及烏喙骨三骨塊相接處有一凹陷,即肩臼,游離的前肢即關節於肩臼處。
爬行動物的肩帶
爬行動物肩帶中肩胛骨與烏喙骨各類中都有,最為穩定。膜原骨除鎖骨外,另有間鎖骨。間鎖骨是早期四足動物新出現的一塊膜原骨,位於腹中線上。大多數爬行動物都有間鎖骨,一直保存到原始哺乳動物。
魚綱動物的肩帶
軟骨魚類的肩帶呈半環形,緊位於咽顱的後方,橫列在身體腹面,不與頭骨或脊柱直接關聯,稱為肩胛烏喙骨,其兩側各有一關節面與胸鰭關節,稱肩臼。
硬骨魚類肩帶位置靠前,由伸向背面的肩胛骨、腹面的烏喙骨、上匙骨、匙骨等組成,並通過上匙骨與頭骨的顳骨相癒合,使頭、肩帶、軀幹形成一個穩定支架,增加了游泳運動的力度,是硬骨魚所持有的。
鳥綱動物的肩帶
鳥綱動物由於適應飛翔生活,肩帶的形態變化較大。肩胛骨為狹長的骨片,位於肋骨的背面,烏喙骨粗大,牢固地與胸骨相連。鎖骨細長,兩側鎖骨呈“V”字形連合,叫叉骨或願骨,富有彈性,阻礙在鼓翼時左右烏喙骨的靠攏,起著橫木的作用。失去飛翔能力的平胸鳥類(如鴕鳥),鎖骨退化,成為烏喙骨的一部分。
哺乳綱動物的肩帶
哺乳綱動物中最低等的單孔目動物(如鴨嘴獸)仍保留著爬行動物的肩帶樣式:軟骨原骨包括烏喙骨、前烏喙骨和肩胛骨;膜原骨包括鎖骨和間鎖骨。有胎盤類哺乳動物,烏喙骨退化成一喙突,附著於肩胛骨上,前烏喙骨和間鎖骨皆已消失。鎖骨的變化較大,在兔、犬、馬等動物,鎖骨退化或消失。兔的鎖骨退化成一小細骨條,埋於肩部肌肉中,僅以韌帶一端連胸骨,另一端連線肱骨。鎖骨的存在與否和運動方式有密切關係。一般來說,善於跳躍、奔跑的哺乳動物,鎖骨大多數退化;而前肢具有多樣性活動的哺乳動物,包括用前肢掘土(如鼴鼠)、飛翔(如蝙蝠)和攀緣(如猴)的種類,鎖骨較發達,這樣的前肢在多樣性活動中具有更大的堅固性。
腰帶
腰帶的演化
在結構上較前肢的肩帶具有更大的堅固性。在脊椎動物各綱中,腰帶的變化較小,全是軟骨原骨,無膜原骨加入。魚的腰帶不和脊柱關連,作用很小,現代四足動物的腰帶則與脊柱相連,成為脊柱與後肢附肢之間的橋樑,起著支持身體的作用。
腰帶的原始類型見於軟骨魚(如鯊魚),這僅是一條橫貫軀幹後部的軟骨,稱坐恥骨棒。棒的兩端微向上突起,稱髂骨突起,可以認為是髂骨的雛型。這樣的腰帶並未和脊柱連線,這是和魚的偶鰭不承擔體重相關的。硬骨魚(如鯉魚)的腰帶很簡單,僅由一對無名骨組成,其內緣後方有小部與對側癒合。
脊椎動物的腰帶
脊椎動物由水生髮展到陸生,四肢需承擔體重,與此相關,腰帶骨與脊椎相連線,這又促使脊柱上有薦椎的分化。在早期的兩棲動物化石中腰帶也有與脊柱不連線的,因此也沒有薦椎的分化。在較後期的堅頭類化石中,可以看出,腰帶向背面延伸,終至與脊往相連線。由此可見,腰帶和脊柱的相連線、薦椎的分化都是和動物由水上陸後,四肢負荷身體重量直接相關的。
兩棲以上動物腰帶
兩棲動物以上的各類動物的腰帶
兩棲動物以上的各類動物的腰帶每側是由髂骨、坐骨和恥骨組成。三骨塊相會合處有一凹,叫髖臼,在此處與後肢的股骨相關節。腰帶的骨塊可以和前肢的肩帶作一對比:髂骨相當於肩胛骨,坐骨相當於烏喙骨,恥骨相當於前烏喙骨。和肩帶不同的是,腰帶的3對骨塊極富保守性,只要腰帶不退化,3對骨塊總是共同出現,這與腰帶所擔負的支持功能和與脊柱的關係是相聯繫的。
蛙或蟾蜍的腰帶已具有3對骨片所組成的典型結構。但蛙類適應於跳躍生活,髂骨特別加長,前端與薦椎的橫突相連,後端與對側的髂骨相連線。
爬行動物腰帶
爬行動物的腰帶也是由髂骨、坐骨和恥骨合成。和兩棲動物不同的是,在兩棲動物中,左右恥骨與坐骨是全部癒合的,而爬行動物的恥骨和坐骨之間分開,形成一個大孔,叫恥坐孔,左右恥骨在中線處結合,叫恥骨連合,左右坐骨結合,叫坐骨連合。這樣的腰帶結構可以減輕重量,而支持的力量並不減弱。龜鱉和鱷腰帶上有大的閉孔,閉孔是由恥坐孔和閉孔神經孔癒合而成,一直到哺乳動物都保留此孔。
在古代爬行動物的分類上,腰帶的結構占有重要地位。根據腰帶的不同,恐龍可分為兩大類,即蜥龍類和鳥龍類。蜥龍類腰帶屬三放型,即髂骨前後走向,恥骨向前下方、坐骨向後下方延伸;鳥龍類腰帶屬四放型,即髂骨前後走向,恥骨和坐骨一併向後延伸,另有恥骨前突伸向前方。四放型的腰帶和鳥類的腰帶近似。
魚綱動物腰帶
軟骨魚類腰帶位於泄殖腔前方,為一橫列的坐恥骨,兩端通過關節面與腹鰭骨關連。不與中軸骨直接相連。
硬骨魚類腰帶簡單,由一對無名骨構成。腰帶不直接與脊柱相連。
鳥綱動物腰帶
鳥類的腰帶具有獨特之處:髂骨、坐骨和恥骨3骨癒合為一塊,並和脊柱的腰薦部(綜合薦骨)癒合在一起,形成一個大而癒合的骨盆,這就增加了它的堅固性,成為後肢強有力的支持者。髂骨為一長大的薄骨片,位於背部,其下後方連線坐骨,兩者之間有髂坐骨孔。恥骨細長,伸延於坐骨的腹緣,兩者之間形成一裂縫狀的閉孔,但3骨的界線在成體中不能辨認。左右恥骨在腹中線處未癒合,構成“開放式骨盆”,這是和鳥類產出硬殼的大卵有關。產卵時,恥骨間距增大,局部去鈣變軟。
哺乳動物腰帶
哺乳動物腰帶的3對骨片癒合成一對無名骨或稱髖骨。左右無名骨與薦椎組成骨盆。在腹側正中線處,兩側的無名骨相結合形成骨盆合縫。雌獸在懷孕時,骨盆合縫之間韌帶變軟,使骨盆腔變大,以利於胎兒的產出。單孔類和有袋類在每一恥骨的前緣伸出一上恥骨,或稱袋骨,有袋類用以支持腹部的育兒袋。水棲獸類,如鯨和海牛的後肢退化,腰帶也很退化,僅殘留一對小骨片。
