
基因轉變
正文
在染色體倍性不發生變化或不存在重複基因的情況下,子囊菌的四分體中所出現的基因不規則分離現象、四分體是子囊菌減數分裂的產物,在子囊中常以一定的順序排列(見連鎖和交換)。粗糙脈孢菌 (Neurospora crassa) 所產生的八個子囊孢子中,每一對孢子都是一個四分體再經一次有絲分裂後生成的兩個細胞。一般說來,它們的基因型是完全相同的(見連鎖和交換)。正常的分離方式是 4:4;5:3、3:5或6:2、2:6或3:1、1:3都屬於不規則分離(圖1)。產生四個孢子的子囊菌例如啤酒酵母(Saccharomyces cerevisiae)中1:3或3:1的分離方式都屬於不規則分離。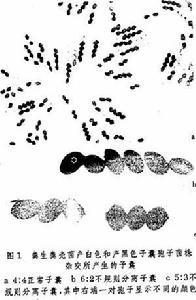 基因轉變
基因轉變基因轉變一般只涉及單個基因,可是一個基因內部的不同突變位點可以分別或同時發生基因轉變,同時發生基因轉變的現象稱為共轉變。共轉變可以發生在相距1000個鹼基對甚或更遠的突變位點之間;同一基因內部的各個突變位點的轉變頻率常從基因的一端向另一端遞減。染色體上基因轉變頻率呈現這種極性現象的小區稱為極化子。
任何一個說明轉變機制的理論必須能夠解釋基因轉變往往伴隨著兩旁基因的重組,以及一對子囊孢子中的兩個孢子有時並不相同這兩個事實。為了說明這些現象,一些學者曾提出不同的模型,其中1964年由美國學者R.霍利代所提出的霍利代模型受到了廣泛的注意(圖2)。它包括:①聯會(這裡只畫出四個染色單體中的兩個);②兩個染色單體雙鏈 DNA分子中的各一個單鏈為核酸內切酶切斷;③單鏈游離端移動;④游離端交換位置;⑤單鏈連線成為半交叉;⑥⑦半交叉位置移動;⑧兩臂鏇轉後成為⑨中空的十字構型;⑩左:左右兩個單鏈切斷,右:上下兩個單鏈切斷;進行DNA修補合成;左:出現中間包含雜合雙鏈部分的兩個染色單體雙鏈DNA分子,兩旁的基因沒有發生重組;右:同左,但兩旁基因發生了重組。
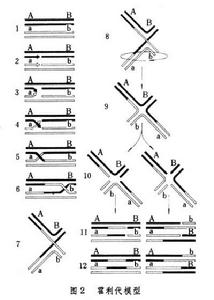 基因轉變
基因轉變兩個雜合雙鏈可以都被校正或者都發生減數後分離,也可以一個被校正而另一個發生減數後分離,而且在校正過程中被切除的可以是這一個或那一個單鏈,因此基因轉變的結果可以導致多種不規則分離現象。
霍利代模型的一個有力的證據是在電鏡照相中觀察到大腸桿菌質粒 ColEl具有圖2中9那樣的DNA 分子圖象。除了力求證實這一模型的實驗工作以外,近年來還出現一些有關這一模型的修改。這一模型不一定完全正確,可是不失為研究交換機制的一個良好的出發點。
配圖
 基因轉變
基因轉變