
簡介
 嘔吐毒素
嘔吐毒素脫氧雪腐鐮刀菌烯醇(deoxynivalenol,DON)又稱去氧瓜萎鐮菌醇(DeoxynivalenolDON)、嘔吐毒素素(vomitoxin),由禾穀鐮刀菌產生,常出現在玉米(穗腐病)小麥和大麥(赤霉病)上。黴菌通常喜好濕冷氣候,菌呈白色或紅色。
解釋
 建議限量
建議限量毒性效應
強制標準
建議限量
理化性質
 DON-BSA偶聯物紅外光譜圖
DON-BSA偶聯物紅外光譜圖DON化學結構為3α,7α,15-三羥基-12,13-環氧單端孢霉-9烯-8酮,屬於B型TCH。DON分子式為
C15H20O6,相對分子質量為296.32。DON純品為白色針狀結晶,熔點為151 ~153℃(醋酸乙基石油)。不飽和酮基的存在使得其在短波紫外下有吸收峰,但與許多其它物質在此處的紫外吸收峰相重疊,屬非特徵性的。紫外輻射下不顯螢光。DON易溶於極性的溶劑如水、甲醇、乙醇、乙腈、丙酮和乙酸乙酯,不溶於正已烷、丁醇、石油醚。DON耐熱、耐壓,在弱酸中部不分解,研究表明,DON在食品加工中,烘焙溫度210℃,油煎溫度140℃、或煮沸,只能破壞50%。加鹼、高壓以及熱蒸汽的處理可以破壞其部分毒力,有研究結果顯示在高壓熱蒸汽作用下可以使其完全失活。在pH=4時,DON在100和120℃下加熱60min其化學結構均不被破壞,170℃加熱60min僅少量被破壞;在pH=7時,在100和120℃下加熱60min仍很穩定,170℃加熱15min部分被破壞;在pH=10時,100℃下加熱60min部分被破壞,120℃下加熱30min和170℃下加熱15min完全被破壞。DON在甲醇中不穩定,22天后被轉化為其它產物。DON的耐藏力也很強,病麥經四年的貯藏,其中的DON仍能保留其原有的毒性。嘔吐毒素(DON)屬於小分子半抗原,有免疫反應性,無免疫原性,只有將其與大分子載體蛋白偶聯,才能作為免疫原,這種人為製備的小分子半抗原與蛋白質的偶聯物稱為人工抗原。可用的載體蛋白有牛血清白蛋白(BSA)、兔血清白蛋白(RSA)、牛甲狀腺球蛋白(BTG)以及雞卵清白蛋白(OVA)等,其中以BSA、OVA最為常用。
人工抗原圖冊參考資料。
 嘔吐毒素紫外光譜
嘔吐毒素紫外光譜蛋白質和半抗原的結合是通過游離的氨基、羧基、酚基、巰基、咪唑基、吲哚基和胍基等活性基團的縮合。連線的方法有物理法和化學法,物理吸附的載體有澱粉、聚乙烯吡咯烷酮、硫酸葡聚糖、羧甲基纖維素等,是通過電荷和微孔吸附半抗原。化學法是利用功能基團把半抗原連線到載體上,常用的方法有戊二醛法、碳二亞胺法、活潑酯法、亞胺酸酯法和鹵代硝基苯法等。半抗原與載體連線時應該注意:帶游離氨基或游離羧
基以及兩種基團都有的半抗原,羧基可用混合酸酐法和碳化二亞胺法與載體氨基形成穩定的肽鍵。同樣,帶氨基的半抗原則可與載體羧基縮合;帶有羥基、酮基、醛基的半抗原,如醇、酚、多糖、核酸以及淄族激素等,它們都不能直接與載體蛋白相連線,需要用化學法在半抗原上引入羧基後才能與載體相連線;芳香族半抗原由於環上帶有羧基,它鄰位上的氫很活潑,極易取代。根據嘔吐毒素本身的結構特性,需要用化學方法在嘔吐毒素上引入羧基後才能與載體相連,然後套用碳二亞胺法合成完全抗原,該法可分為兩種,一種是在有機相中進行反應的二環已基碳二亞胺法(dicylochexylcarbodiimide,簡稱DCC),另一種是在水相中進行反應的對乙基-N,N-二甲基丙基碳二亞胺法(3-mercaptopropionicacid,1-ethyl-3-(3-dimethylaminopropy1)carbodiimide,簡稱EDC)。DCC法需要在嚴格的無水條件下進行反應,所用的有機溶劑需預先脫水處理,需要乾燥的反應環境;EDC法可以直接在水相中進行,但EDC需要超低溫(-20℃)避光保存。
毒性
生物體特徵
嘔吐毒素對人和動物均有很強的毒性,能引起人和動物嘔吐、腹瀉、皮膚刺激、拒食、神經紊亂、流產、死胎等,豬是對嘔吐毒素最敏感的動物,家禽次之,反芻動物由於瘤胃微生物的作用,耐受力最強。對生長肥育豬而言,含有12~14g/t嘔吐毒素的飼料飼餵後10~20min即會出現嘔吐、不正常的焦慮和磨牙現象,含毒量19g/t以上即完全拒食。研究表明,不同種類動物的LD50(mg/kg)為:小鼠經口46.8腹腔注射70.0;新生大鼠經口7.3;大鼠徑口7.3;北京雛鴨皮下注射27.0;而致吐劑量為:北京雛鴨皮下注射10.0;狗皮下注射0.1經口0.1;豬腹腔注射0.05;鴿子經口10.0。
幾乎所有的單端孢霉烯族毒素都有可引起鴨雛、貓、狗、豬、鴿子等動物的嘔吐。豬對DON的致吐作用最敏感,約為其他動物的100-200倍。DON還可引起拒食反應。致嘔吐作用的機理除了DON對黏膜的刺激作用外,同時也有對中樞神經的作用。曾有人以含DON毒素18-20mg/kg的赤霉病小麥,按0.5-50﹪的不同比例混入飼料中,餵養大鼠18個月,發現高比例組動物睪丸、子宮、肝、腎臟受到一定的損害。
DON化學性能非常穩定,一般不會在加工、儲存以及烹調過程中破壞,在實驗室條件下可長期貯存保持毒力不變,有較強的熱抵抗力,121℃高壓加熱25min僅有少量破壞。酸性環境不影響其毒力,但是加鹼或高壓處理可破壞部分毒素含有嘔吐毒素穀物類加工產品,在加工過程中,由於技術限制不可能完全去除,人類食用含有一定量嘔吐毒素糧油食品後,會出現胃部不適、眩暈、腹脹、頭痛、噁心、嘔吐,手足發麻、全身乏力、顏面潮紅、以及食物中毒性白細胞缺乏症。症狀嚴重者可見呼吸、脈搏、體溫及血壓的波動,四肢發軟、步態不穩、形似醉酒,故有的地方稱其為“醉谷病”。
嘔吐毒素的產毒菌株適宜在陰涼、潮濕的條件下生長。廣泛存在於大麥、小麥、玉米和燕麥中。不同動物對嘔吐毒素的敏感程度不同,豬是對嘔吐毒素最敏感的動物,斷奶仔豬尤其敏感。飼料中0.3~0.5mg/kg的微量毒素就會引起豬拒食、生長下降以及對傳染病的抵抗力下降,飼料中含1mg/kg以上的嘔吐毒素時可引起豬拒食、嗜睡、生長嚴重受阻、體增重減慢、免疫機能減退、肌肉協調性喪失以及嘔吐等症狀。此外,嘔吐毒素可在人和動物體內蓄積,具有致畸性、神經毒性、胚胎毒性和免疫抑制作用。
 嘔吐毒素對微血管影響
嘔吐毒素對微血管影響嘔吐毒素主要在胃腸道吸收,經血液循環進入肝臟代謝後經腎臟隨尿液排出,造成各器官的廣泛損傷。嘔吐毒素在人體中的半衰期為2~4h,代謝迅速,8h後就檢測不到嘔吐毒素和它的代謝物。
 嘔吐毒素對肝功能影響SEM圖
嘔吐毒素對肝功能影響SEM圖嘔吐毒素可引起雛鴨、豬、貓、狗、鴿子等動物的嘔吐反應,嚴重者可造成死亡。嘔吐毒素的急性毒性與動物的種屬、年齡、性別、染毒途徑有關,雄性動物對毒素比較敏感。急性中毒的動物主要表現為站立不穩、反應遲鈍、豎毛、食慾下降、嘔吐等,還可引起動物的拒食反應,其中豬對嘔吐毒素最為敏感。嘔吐毒素能引起豬食慾減退或廢絕,嘔吐,體重下降,流產,死胎和弱仔,抑制免疫機能和降低機體抵抗力。對生長肥育豬而言,含有14mg/kg嘔吐毒素的飼料飼餵後10~20min即會出現嘔吐、不正常的焦慮和磨牙現象,嘔吐現象僅發生在第1天。而且嘔吐毒素含量在0~14mg/kg的試驗中,飼糧中每增加1mg/kg嘔吐毒素,生長肥育豬的採食量減少6%,在含毒量10mg/kg以上即完全拒食。對於嘔吐毒素的慢性毒性世界研究都比較少。有人用大鼠進行了為期2年的染毒試驗,嘔吐毒素的濃度為0、1、5、10mg/kg,雄性、雌性大鼠各分為4組,試驗結束後發現,各組動物均未見死亡,動物體重增加與染毒劑量呈負相關。雌性大鼠的血漿中IgA、IgG濃度較對照組飼料中嘔吐毒素的危害與防治增高,生化指標、血液學指標也可見明顯異常。病理學檢查還發現有肝臟腫瘤、肝臟損害。
長期攝入被DON污染的病麥或粗毒素對動物有一定的毒性,可影響動物的生長、繁殖率和子代的存活率,甚至可使生長停滯,形成“僵鼠”,並產生對心、肝、腎的傷害,嚴重時損害造血系統造成死亡。但不同種屬、性別的動物對DON的敏感程度不一,豬是最敏感的動物,反芻動物比其它動物受的影響小,奶牛的健康狀況、產奶量、攝食量幾乎不受影響,牛奶中也檢測不到DON及其脫氧產物。
在很多物種上已經得到證實,DON可以通過胎盤轉移到胎兒。懷孕母豬感染DON後在胎兒的血漿、肝和腎中均檢測到DON。用DON純品灌餵Swis小鼠,引起明顯的胚胎毒作用和骨骼畸形:DON在10~15mg/kg劑量時胎兒100%吸收,在5mg/kg的劑量時胎兒吸收率為80%。在大鼠妊娠期5~19d中飼餵5mg/kg的DON會降低胎兒的發育,母鼠採食量和平均日增質量都顯著降低,胎兒質量、母體子宮質量和頭臀長降低,胸骨、趾骨、背骨和椎骨等骨化能力下降,這些可能是由於母體毒性和胎兒大小減小造成的。在齧齒動物上的致畸和繁殖試驗證實DON影響大鼠的體質量增長,導致吸收胎增多、骨骼形成不良及仔鼠出生後存活率的降低。剛斷奶的幼鼠除了在血漿和體組織中檢測到DON質量濃度明顯高於成年鼠以外,DON誘導幼鼠脾和肺中產生的IL-6和TNF-α質量濃度也比成年鼠的高,持續的時間更長,且脾中IL-6、IL-1β和TNF-α的mRNA表達量比成年鼠高出2~3倍,這些均表明幼鼠對DON引起的多種影響的反應比成年鼠敏感,所以DON對幼鼠機體組織造成的負荷更大。Vesely等研究了DON對於3日齡雞胚的毒性作用,發現其胚胎毒性劑量範圍很窄(1~3mg/kg),DON致使試驗組雞胚頭部畸形,身體發育畸形,畸形率明顯高於對照組。
毒素污染分布
有調查顯示,中國飼料和原料黴菌毒素污染超標的比例為60%~70%,其中嘔吐毒素的超標比例接近70%,在被檢的玉米、麩皮和DDGS樣品中嘔吐毒素的檢出率均高達90%以上,分別為92.9%、92.3%和100%。豆粕的檢出率較低為54.5%。麩皮和豆粕中嘔吐毒素未見超標,其平均含量分別為0.44mg/kg和0.05mg/kg,屬於輕度污染。玉米中嘔吐毒素的超標率為57.1%,毒素平均含量為1.01mg/kg,其最高含量為2.13mg/kg,屬於中度污染。DDGS中嘔吐毒素的超標率為88.2%,毒素平均含量為1.36mg/kg,最低含量為0.85mg/kg,最高含量為1.72mg/kg,屬於中度污染。玉米和DDGS中嘔吐毒素的平均含量極顯著高於麩皮和豆粕(P<0.01),DDGS中嘔吐毒素的平均含量顯著高於玉米(P<0.05),麩皮中嘔吐毒素的平均含量極顯著高於豆粕(P<0.01)。被檢配合飼料中嘔吐毒素的檢出率為97.4%,仔豬料、中豬料、妊娠母豬料和哺乳母豬料的檢出率高達100%。其中,乳豬料嘔吐毒素未見超標,其平均含量為0.28mg/kg,顯著低於其他種類全價料(P<0.05),屬於輕度污染。仔豬料、中豬料、大豬料、妊娠母豬料和哺乳母豬料均有不同程度的超標,超標率分別為14.3%、14.3%、22.2%、10.0%和23.1%,其最高含量分別為2.31、1.14、1.44、1.45mg/kg和1.52mg/kg,屬於中度污染。
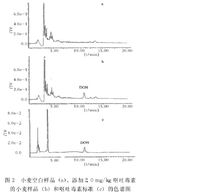 小麥麵粉中嘔吐毒素色譜圖
小麥麵粉中嘔吐毒素色譜圖小麥麵粉中嘔吐毒素色譜圖圖冊參考資料。
DON的污染廣泛存在於全球各國,中國、日本、美國、前蘇聯、南非等均有發現。DON主要污染小麥、大麥、燕麥、玉米等穀類作物,也污染糧食製品,如麵包、餅乾、麥制點心等。另外,在動物的奶、蛋中均有發現DON殘留。DON對於糧穀類的污染狀況與產毒菌株、溫度、濕度、通風、日照等因素有關。小麥赤霉病主要分布在潮濕的溫帶地區,中國大部分地區又恰恰處於這一地帶,這是中國DON污染較嚴重的原因之一,在多雨年份DON的污染狀況則更為嚴重,赤霉病麥中毒也是中國最主要的真菌性食物中毒之一,這也越來越引起了人們的重視。
 四種鐮刀菌產生嘔吐毒素情況
四種鐮刀菌產生嘔吐毒素情況DON毒性較低,但它最易出現,廣泛存在於溫熱帶地區的飼料中,因此在穀物中DON中毒發病率最高。例如:在日本,小麥和大麥中的DON濃度為40ppm;在加拿大,安大略地區白色冬小麥中的DON濃度為8.5ppm。對一些亞洲國家的未加工的農產品和加工過的飼料樣品進行檢測時發現,中國的飼料樣本中,含有一定量的嘔吐毒素。超過70%的樣本被嘔吐毒素污染。2000年的樣本中嘔吐毒素的最高含量達到4582μg/kg。2003年中國配合飼料樣品中嘔吐毒素的陽性檢出率為100%,平均含量為600μg/kg;據估計,2005年中國飼料原料的嘔吐毒素檢出率為100%,對人類的健康產生威脅。美國食品及藥物管理局(FDA)規定食物中嘔吐毒素的安全標準是1000μg/kg,嘔吐毒素的含量超過l000μg/kg就會對人及一些動物健康產生損害。
嘔吐毒素對中國穀物類原料的污染相當普遍,其次是油籽類原料。從內蒙、寧夏、黑龍江、遼寧、湖南、湖北、河北、廣東等省市採集31份玉米、21份全價飼料、27份植物蛋白飼料。被檢的玉米樣本中嘔吐毒素檢出率達100%,平均含量達820μg/kg;被檢全價料中嘔吐毒素的檢出率達100%,平均含量為1020μg/kg;蛋白質飼料中嘔吐毒素檢出率達87%,平均含量為240μg/kg。據顧薇2003年研究發現,從中國送來的樣本中,超過70%的樣本被嘔吐毒素污染,1998-1999年、2000年和2001年,嘔吐毒素的平均濃度分別為548,607和378μg/kg}2000年的樣本中嘔吐毒素的最高含量達到4582μg/kg。據奧特奇公司研究結果,2005年中國飼料原料嘔吐毒素檢出率為100%,超標率27.3%(最高為89.81ppb)。
 四種鐮刀菌產生嘔吐毒素電泳圖
四種鐮刀菌產生嘔吐毒素電泳圖鐮刀菌屬黴菌屬於田間黴菌,野外菌株,通常在作物生長期間就被污染,其最適生長溫度為5~25℃,由於鐮刀菌屬黴菌的上述特點,嘔吐毒素的分布與對飼料原料的污染也呈現出一些特點:1)鐮刀類黴菌主要在田間
污染穀物類原料作物和油籽類原料作物。嘔吐毒素則是在剛收穫後的穀物嘔吐毒素污染較為嚴重。2)嘔吐毒素對中國穀物類原料的污染相當普遍,其次是油籽類原料。3)嘔吐毒素的污染程度存在地區和年份的區別。由於溫度、濕度等的差別,不同地區、同一季節收穫的玉米所帶菌屬有較大差別,同一地區、不同季節、不同年份的玉米所帶菌屬也不一樣。北方地區的玉米(如河南、山西等地的部分玉米)以鐮刀菌為主要菌屬,而東北玉米以圓弧青黴為主要菌屬,鐮刀菌次之。所以,華北地區的部分玉米嘔吐毒素的污染較重一些,而東北玉米相對較輕一些。就年份而言,與收穫時的降雨量有關。如2003年,華北部分地區,由於下雨太多,造成部分玉米嘔吐毒素污染嚴重,少數玉米用於生產飼料,嚴重影響飼料的適口性,導致飼料退貨。而2004年因收穫時天氣較好,嘔吐毒素沒有達到危害的程度。2005年的新玉米,部分玉米又有不同程度的超標。
細胞毒性
DON具有很強的細胞毒性,它對於原核細胞、真核細胞、植物細胞、腫瘤細胞等均具有明顯的毒性作用。Prelusky等發現,豬餵給嘔吐毒素,能提高腦中5-羥色胺(serotonin)和5-羥引哚乙酸(5-Hydroxyindoleaceticacid)的濃度,對腦神經產生麻痹作用。而且,即使是低劑量的DON就能引起豬腦中5-羥引哚乙酸水平的提高。Rizzo等發現不同濃度DON對於大鼠紅細胞的溶血作用,其溶血作用有一個閾值,低於此濃度紅細胞不會發生溶血反應。DON可能通過3種不同的方式對原核細胞產生毒性作用:(1)通過滲透磷脂雙層,作用於亞細胞水平;(2)通過與細胞膜相互作用;(3)通過自由基介導的脂質過氧化作用。DON可能以一種以上的方式同時發揮作用。Parent-Massin等研究發現,DON可作用於骨髓造血細胞而產生細胞毒性;還可作用於T細胞、B細胞、IgA+細胞而產生免疫毒性作用,抑制或增加細胞程式死亡。其免疫毒性作用依賴於DON的濃度、淋巴細胞亞型、組織來源和糖皮質激素。還有研究指出,DON可以抑制胸腺細胞的蛋白質合成。多數研究都表明DON具有胚胎毒性和致畸作用。動物試驗表明,長期小劑量餵飼含毒素飼料,可以誘發不同器官的腫瘤。Zhou等(2003、1997)經口給予小鼠5mg/kg和25mg/kg嘔吐毒素,結果致炎細胞因子產生IL-1β、IL-6和TNF-α的mRNA的表達;Th1細胞IFN-γ和IL-2mRNA、Th2細胞IL-4和IL-10mRNA的表達都明顯升高。Thuvander等發現,嘔吐毒素在高濃度可劑量依賴抑制體外培養的人外周淋巴細胞增殖及免疫球蛋白的產生,而低濃度則使免疫球蛋白升高。一次性腹腔注射嘔吐毒素,12h後可觀察到小鼠胸腺發生凋亡的細胞明顯增多,增殖活性受到抑制,而且隨著嘔吐毒素濃度的升高,這種作用越明顯。
免疫毒性
研究表明,DON可能對免疫系統有影響。DON既是一種免疫抑制劑,又是一種免疫促進劑,其作用與劑量有關。在體內DON可以抑制對病原體的免疫應答,同時又可以誘發自身的免疫反應。DON引起實驗動物免疫系統的疾病與人類lgA腎病及其相似,同時DON還可以誘發輔助性T細胞超誘導產物-細胞因子的產生,激活巨噬細胞、T細胞產生炎症前細胞因子。
動物毒力學試驗研究表明:不同畜種的動物對DON的敏感程度按順序是:豬>小鼠>大鼠>家禽和反芻動物。DON對人畜會產生廣泛的毒性效應,低劑量攝入會造成食慾下降、生長緩慢、發育不良和病死率升高等現象;大量或長期攝入會發生急性中毒、嘔吐、直腸出血和腹瀉,與腸道炎症極其相似。嘔吐毒素對動物機體毒性主要分為免疫毒性和細胞毒性。DON在低質量濃度時能誘導輔助性T細胞超誘導產物—炎性細胞因子和趨化因子在巨噬細胞中的表達。高質量濃度DON可抑制免疫,誘導免疫細胞凋亡,抑制其增殖,同時還影響免疫細胞因子的分泌、減弱免疫應答、抑制淋巴細胞增殖、吞噬作用和巨噬細胞的殺菌作用。
三致作用
世界對此研究結果不太一致,但多數研究結果均表明DON具有三致(即致癌、致畸、致突變)作用,可能是一種潛在的致癌物質。DON的誘癌實驗尚未成功,世界也無其致癌作用的明確研究結論,因此其致癌作用尚無定論。但流行病學研究發現,DON與食管癌的發病率呈正相關。有動物實驗表明,長時間小劑量餵食被嘔吐毒素污染的飼料,可以誘發不同器官的腫瘤。Iserson等進行的DON慢性毒性實驗發現,實驗組動物發生肝癌。Lamber LA等用大鼠進行了DON兩階段皮膚誘癌、促癌實驗,結果表明DON不是一種致癌劑或誘癌劑,但皮膚組織學發現DON誘發瀰漫性磷狀上皮增生。Vesely等研究了DON對三日齡雞胚的毒性作用,經8天孵化後發現DON的毒性作用劑量範圍很窄,為1-3mg/kg體重,實驗組雞胚有頭部畸形、身體發育畸形,畸形率明顯高於對照組。
毒理學
對免疫細胞的影響
胸腺是T細胞發育、分化、選擇和成熟的場所。原胸腺細胞經過前胸腺細胞、CD4-CD8-雙陰性細胞。CD4+CD8+雙陽性細胞和CD4+CD8-或CD4-CD8+2個單陽性胸腺細胞亞群這5個階段,發育分化為成熟的T細胞。因此T細胞在胸腺整個發育過程中會先後經歷陽性選擇和陰性選擇,經過陽性選擇的單陽性胸腺細胞既可識別異己抗原又可識別自身抗原,但識別自身抗原這一特性對機體是有害的。只有與自身抗原親和力低和不識別自身抗原的胸腺細胞才會經過陰性選擇後進一步分化為成熟的T細胞,反之就會發生細胞凋亡。所以T細胞在胸腺活化時會引發胸腺細胞的陰性選擇和凋亡,而且在T細胞活化或胸腺細胞陰性選擇期間,DON促使抑制免疫的基因表達升高,所以胸腺對DON有高敏感性。在小鼠口服5、10和25mgDON/kg體質量各3、6和24h後,Sandra分析基因時證實在前胸腺細胞階段DON選擇性的使抑制免疫的基因(抑制線粒體、核糖體或蛋白質合成的基因)高度表達,也證明了在胸腺細胞發育成T細胞過程中雙陽性CD4+CD8+階段對DON最敏感,高質量濃度DON能明顯誘導胸腺細胞凋亡,抑制其增殖。
但是,在T細胞活化反應期間,用低質量濃度DON刺激胸腺細胞3h,很多基因,如:鈣依賴基因和核內轉錄因子(NFkB)的靶基因被刺激表達,誘發內質網鈣儲存的消耗,活化NFkB,導致大量鈣離子依靠鈣轉運蛋白流過細胞膜,胞內鈣質量濃度增加,激活鈣調磷酸酶,使活化T細胞核因子(NFAT)去磷酸化,DON誘導NFAT轉至胸腺細胞核內,活化更多基因,促進T細胞活化和增殖。
巨噬細胞在動物機體的免疫防禦中起著關鍵作用,作為機體免疫應答的組成部分,構成免疫防禦的第一道防線,同時也參與機體的獲得性免疫。巨噬細胞被活化後產生不同的炎性因子,並表達一些特定的細胞表面受體或白細胞分化抗原(CD)。研究表明:低質量濃度DON可提高個別炎性因子的分泌和產生,高質量濃度DON則誘導巨噬細胞的凋亡。腫瘤壞死因子(TNF)-α是巨噬細胞活化的主要因子,依賴劑量提高細胞表面受體(CD14、CD54和CD119)和人類白細胞抗原(HLA-DP/DQ/DR)的表達,而DON被證實不論質量濃度高低均會干擾TNF-α活化巨噬細胞的過程,起到免疫毒性的效果。DON在細胞因子受體和兩面神激酶(JAK)水平上作用於細胞因子信號抑制因子(SOCS),抑制細胞因子受體(TNF-α受體)的信號轉導,進而抑制多種細胞因子共用的信號轉導途徑—酪氨酸激酶/信號轉導子和轉錄激活子(JAK/STAT)的信號轉導,干擾TNF-α刺激巨噬細胞活化的過程。鼠巨噬細胞中脂多糖(LPS)誘導一氧化氮聚合酶(iNOS)的表達,干擾素(IFN)-β是該誘導過程不可缺少的信號,IFN-β的表達引起STAT、干擾素調控蛋白和iNOS表達。DON還可以抑制iNOS和IFN-β的激活和表達,直接或間接抑制NO和IFN-β的產生。
對細胞因子的影響
有人發現DON除了引起鼠的胸腺、脾、骨髓細胞和集合淋巴結的凋亡,還顯著影響細胞因子的分泌,可誘導淋巴組織表達TNF-α、白細胞介素(IL)-1β和IL-6等促炎症細胞因子。豬體內試驗表明:DON可誘導巨噬細胞產生細胞生長抑制素,尤其是TNF-α和IL-1β(Doll等,2009)。DON在100或250ng/mL的質量濃度時可迅速提高鼠單核巨噬細胞白血病-RAW264.7中IFN-γ和IL-6的mRNA穩定性,延長其半衰期(從30min提高至大於3h)。推測DON通過以下3種方式超誘導促炎性細胞因子的表達,提高其穩定性:1)作為一種翻譯抑制劑,使核糖體凝固或群聚在mRNA上以保護mRNA免受細胞質中核糖核酸酶的裂解;2)與TCH誘導的促細胞分裂原活化蛋白激酶(MAPK)活化有關;3)保護富含腺嘌呤尿嘧啶(AU)序列的3′非翻譯區免受內切酶或外切酶的降解,延長IFN-γ和IL-6mRNA的半衰期(Wong等,2001)。DON明顯增加鼠T細胞中IL-2水平,在基因水平上使用轉錄抑制劑發現DON超誘導IL-2mRNA的表達,且在某種程度上提高IL-2mRNA的穩定性;同時也可提高IL-4、IL-5、IL-6和IFN-γ質量濃度。IL-8是機體最主要的炎症因子,對中性粒細胞和CD4+CD8+有明顯趨化作用。質量濃度為250~1000ng/mL的DON能誘發鼠IL-8轉錄調節因子IL-8mRNA和IL-8hnRNA的產生(二者與IL-8蛋白質的表達和P38絲裂原活化蛋白激酶磷酸化是同時發生的),激活MAPKs信號通路,在炎症與細胞凋亡等應激反應中發揮重要作用。
對免疫球蛋白的影響
免疫球蛋白對DON的敏感性受物種、DON質量濃度及中毒時間等因素的影響。已有研究證實DON顯著影響人免疫功能,被DON感染後人淋巴細胞中免疫球蛋白A(IgA)、免疫球蛋白G(IgG)和免疫球蛋白M(IgM)分泌顯著降低。Goyarts發現豬血液上清液中IgA、IgG和IgM的質量濃度幾乎不受DON的影響,慢性中毒時DON也不顯著影響血清中IgA、IgG和IgM的質量濃度,只有在急性中毒時發現血清中IgG和IgM的質量濃度顯著提高,但血清中IgA不受影響。不過給豬飼餵含5.5mg/kgDON的日糧21d後檢測到血清中IgA和IgM的質量濃度顯著提高,IgG質量濃度沒有受到影響。給大鼠飼餵含6.25mg/kgDON的日糧1周后檢測血清中IgA、IgG和IgM的質量濃度均顯著提高,而Forsell卻發現在大鼠感染DON8周后,依賴劑量下調大鼠血清中IgG和IgM質量濃度的同時上調血清中IgA質量濃度。豬或鼠在長時間受DON感染後,血清中IgA質量濃度的升高引起的疾病與人類IgA腎病極其相似。在細胞水平上,DON不直接影響骨髓或脾中的B細胞分泌IgA,而是會提高脾中T細胞的數量和CD4+/CD4+CD8+的比例,間接影響IgA的分泌。CD4+在DON的脈衝刺激下既可以輔助B細胞分泌IgA,又可提高IL-2、IL-4、IL-5和IL-6分泌蛋白與mRNAs的含量,其中IL-6是輔助IgA分泌的重要因子。在大鼠試驗中也證實DON促進輔助性T細胞因子的產生,加強了lgA分泌時T細胞的輔助作用。在基因水平上,DON通過提高轉錄和mRNA的穩定性2種方式上調內源性環氧酶-2(COX-2)基因的表達,因COX-2可調節DON誘導IL-6基因的表達上調,所以DON間接輔助IgA的分泌。最後DON也可直接誘導鼠中IgA基因表達上調,導致IgA分泌增多,從而大量IgA和IgA免疫複合物在腎小球系膜區沉積,激活血清補體旁路途徑,引起腎損傷。不過在患有過敏症的鼠中DON不增加過敏原特異性引起的免疫球蛋白E(IgE)或IgG1的表達,表明在吸入或攝取DON後不會加重針對過敏原引起的過敏反應。
對卵母細胞的影響
DON誘導淋巴組織表達的TNF-α、IL-1β和IL-6等促炎症細胞因子可損害生育能力,尤其是TNF-α已證實會影響豬和牛卵母細胞的發育。卵母細胞在減數分裂過程中與其周圍的卵丘細胞(濾泡細胞在卵母細胞發育時凸向濾泡腔所形成)形成卵胞質,在卵胞質結構和分子的變化及2種細胞間的相互作用下卵母細胞逐漸成熟。不論卵丘細胞的來源—濾泡細胞的大與小,其對卵母細胞成熟過程的作用都是不會改變的,所以卵丘細胞的擴增和死亡是卵母細胞獲得發育能力的關鍵因素。在豬卵母細胞成熟過程中,用DON感染其卵母卵丘細胞複合體(COCs)後,檢測發現大量卵丘細胞死亡的同時卵母細胞潛在的發育能力下降,由此推測DON對卵母細胞的毒性是通過抑制卵丘細胞增殖和控制卵丘細胞周期間接作用而導致的。減數分裂中紡錘體是在第一次分裂中期形成的,紡錘體形成是微管的動態組成。微管合成的異常和受精後染色體的異常分裂會造成胚胎髮育受到抑制。Schoevers等在DON感染後的COCs中發現,隨著DON質量濃度的增加,到達第二次分裂中期的卵母細胞顯著減少,而且顯微鏡檢測出很多卵母細胞的核染色質和微管發生畸變,說明成熟卵母細胞在中期I到中期II對DON最敏感。當卵母細胞感染高質量濃度(2μmol)DON時,畸變發生在減數分裂紡錘體形成時期—中期I(在此質量濃度下卵丘細胞增殖被完全抑制);若是低質量濃度,畸變更容易發生在末期I或中期II。DON也會在以下3個階段損害卵母細胞發育以降低胚胎髮育:1)減數分裂紡錘體的形成過程中;2)減數分裂紡錘體形成前(因為DON已經在卵母細胞成熟前21h中降低了其發育潛力);3)受精過程中。DON還可通過直接影響動粒蛋白(紡錘體形成的關鍵點)而干擾減數分裂紡錘體形成,使減數分裂停滯在中期I或末期I。
脫毒與防治
嘔吐毒素的污染已引起人們的高度重視,但許多國家的飼料及飼料原料嘔吐毒素限量標準和污染程度判定標準尚不完整,也不統一。如美國制定了飼料用穀物及其副產品(除玉米外)嘔吐毒素允許限量≤5mg/kg,歐盟制定玉米及其副產品允許限量≤1.75mg/kg。中國制定了部分配合飼料(如,豬、禽及犢牛)中嘔吐毒素的限量標準,但未制定飼料原料(除DDGS外)中嘔吐毒素限量標準。
1993年,DON被世界衛生組織(WTO)的癌症研究機構(IARC)劃定為第3類,即“無法分類為對人類有致癌效應”的物質。基於長達2年的低劑量DON小鼠餵養實驗表明DON不具有致癌的危害,得出NOEL(每日100μg/kg)及100倍的安全因子,食品添加劑國際法典委員會JECFA提出了一個不會對人類免疫系統、生長和生殖產生負面影響的DON的PMTDI為1μg/kgb.w.,並認為DON的現有數據不足以建立一個不會導致人類急性疾病暴發的DON水平。委員會還就膳食中DON的攝入量進行了初步評估,結果發現世界上許多地區膳食中DON的攝入量可能會超過此值,但是考慮到在攝入量估計過程中由於DON的含量水平和消費量的不確定性,以及食品加工過程中對DON濃度的不同程度的降低,會使DON攝入量的估計產生極大的不確定性。因此,委員會維持了DON的PMTDI為1μg/kg。
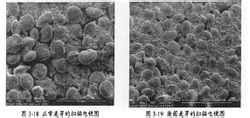 正常與污染麥芽SEM圖
正常與污染麥芽SEM圖嘔吐毒素的防治首先是防止黴菌的產生,而防霉關鍵在於要嚴格控制飼料和原料的水分含量、控制飼料加工過程中的水分和溫度、選育和培養抗黴菌的飼料作物品種、選擇適當的種植或收穫技術、注意飼料產品的包裝、貯存與運輸、添加防霉劑等。但需注意的是使用防霉劑無法去除飼料原料中已存在的黴菌毒素,添加防霉劑只是預防作用,所以飼料的脫毒也是必要的一項措施。飼料及飼料原料發生霉變後,產生的嘔吐毒素(脫氧雪腐鐮刀菌烯醇)是一種無色針狀結晶具有較強的熱抵抗力。因此,可以根據飼料霉變的程度採取不同的方法進行脫毒處理。一般有物理脫毒法、化學脫毒法、酶解法。
物理脫毒法主要是水洗法、剔除法、脫胚去毒法、溶劑提取法、加熱去毒法、輻射法等; 化學脫毒法主要是採用鹼或氧化劑進行處理脫毒。上述的諸多脫毒方法,在實際套用中對於飼料業、養殖業來說均不適用。因其操作不但困難,且大批量的飼料及原料用此類方法是無法進行, 而且經化學脫毒處理後往往會降低飼料的營養品質和適口性。
酶解法主要是選用某些酶,利用其降解作用,使黴菌毒素破壞或降低其毒性。與物理學和化學方法相比,酶的降解處理法對飼料營養成分的損失和影響較少,但因其費用高,效果不穩定製約著該方法的廣泛套用。
吸附法即在飼料里添加黴菌毒素吸附劑進行脫毒的一種方法,這種方法仍是一個比較可行的方法。但在黴菌毒素吸附劑的選用上,應認真考慮。因為吸附劑選用不當,不但起不到吸附黴菌毒素的作用,還會產生副作用。黴菌毒素吸附劑的選用,一般應考慮以下幾方面因素:1)必須具備高吸附能力。原則上選擇產品比表面積在600以上的吸附劑。2)選擇性吸附。理想的黴菌毒素吸附劑應具備只吸附毒素,不吸附營養物質的特點。3)廣譜吸附。因為通常污染原料的黴菌毒素不只一種,所以選擇的黴菌毒素吸附劑一般應對多數黴菌毒素都有效。4)無副作用。
檢測方法
嘔吐毒素的檢測方法有薄層色譜法(TLC)、高效液相色譜法(HPLC)、柱淨化法結合電子捕獲檢測器的氣相色譜法(GC/ECD)、高效液相色譜串聯質譜法(HPLC-MS/MS)、放射性免疫測定法(RIA)等; Casale等則提出了酶聯免疫(ELISA)檢測方法。在中國,魏潤蘊等提出了採用甲醇-水提取,以XAD-4柱淨化,雙向展開的薄層色譜檢測方法及氣相色譜法;郭玉鳳等提出了使用PIB-CI(氯化聚異丁烯)衍生化的液相色譜檢測方法;陽傳和等提出了酶聯免疫吸附的測定方法;張鵬等提出了採用免疫親和柱(IAC)或多功能淨化柱(MFC)淨化結合高效液相色譜法。
色譜法
 嘔吐毒素HPLC譜圖
嘔吐毒素HPLC譜圖薄層色譜法(TLC)是GB/T5009.111-2003採取的穀物及其製品中嘔吐毒素的檢測方法,其檢測限為1mg/kg。適用於穀物(小麥、玉米、大麥等)及其製品(蛋糕、餅乾、麵包等)和中嘔吐毒素的測定。穀物及其製品中的嘔吐毒素經乙腈-水(84:16 V/V)提取、淨化、濃縮和矽膠G薄層展開後,加熱薄層板。在製備薄層板時加入三氯化鋁,使DON在波長365nm紫外光下顯藍色螢光,與標準比較。薄層色譜法TLC雖然操作簡便,曾被廣泛套用,但是,處理樣品時工作量大;在檢測過程中操作人員必須直接接觸標準品,危害到操作人員的身體健康,而且本方法在提取過程中需要使用大量的有機溶劑,這樣就會對周圍環境也會產生不利影響;靈敏度差,提高靈敏度必須改進樣品的提取和淨化方法,改進和提高薄層色譜分析性能,這樣就增加了勞動強度和檢測成本。
嘔吐毒素HPLC譜圖參考資料。
 嘔吐毒素標準品色譜圖
嘔吐毒素標準品色譜圖DON經HPLC分離後可用螢光檢測器(fluorescencedetector,FLD)、紫外檢測器(ultravioletdetector,UV)、二極體陣列檢測儀(diodearraydetector,DAD)檢測。實驗證明,DAD比FLD的效果好:不需衍生;DAD最低檢出限比FLD要低;DAD檢測比FLD回響強。2001年Mateo使用DAD的最低檢出限可達5μg/kg,而FLD的最低檢出限為25μg/kg。穀物中DON的HPLC-UV的最低檢出限可達100~1600μg/kg;HPLC-MS可達1240μg/kg;HPLC-FLD可達20μg/kg。運用氣相色譜檢測DON需要通過DON上的三個羥基使之衍生成為七氟丁醯(heptafluorobutyryl,HFB)、三氟乙醯(trifluoroacetyl,TFA) 或三甲基矽烷(trimethylsilyl,TMS) 化衍生物。
高效液相色譜法(HPLC)和氣相色譜(GC)可以精確地對樣品中的嘔吐毒素進行定性定量分析。測定DON需進行衍生化,通過三甲矽烷衍生物,利用電子捕獲檢測器進行定量分析,操作繁瑣、重現性較差。而且這兩種方法所需色譜儀、檢測器等價格昂貴,而且樣品前處理比較複雜,操作時需要專門的技術人員,不便推廣套用。另外,它們也不適合大批量樣品的檢測。
酶聯免疫吸附測定法
免疫酶技術(EIA)根據抗原抗體反應後是否需要分離結合的與游離的酶標記物而分為均相(homogenous)和異相(heterogenous)兩種類型。在異相法中,又包括液相異相酶免疫測定和固相酶免疫測定二種。固相酶免疫測定也稱為ELISA,是臨床檢驗中套用較廣的免疫測定方法。小分子的ELISA測定法一般有三種競爭分析模式:包被特異性抗體直接競爭法、包被抗原直接競爭法以及間接競爭法(也稱為抑制性測定法)。間接競爭ELISA法的基本步驟是:包被完全抗原——同時加入已知抗體和待測抗原——加入酶標二抗——加入底物顯色液。該法主要套用於未知抗原的檢測。
酶聯免疫吸附法(ELISA)快速、靈敏、準確、可定量、操作簡便、無需貴重儀器設備,且對樣品純度要求不高,特異性強,特別適用於大批量樣品的檢測,發展非常迅速。簡化了樣品預處理和純化過程。但是由於免疫酶的活性非常不穩定,在套用過程中很容易受到操作條件的影響,反應試劑需低溫保存。在實際操作過程中很難達到這一要求,使得免疫酶的活性大大降低,從而影響到結果的準確性。套用時還必須通過多次洗滌,才能使反應產物和游離底物進行分離,影響到結果的重複性,而且檢測時間較長,無法滿足現場快速檢測的要求。
檢測
DON的污染常出現於小麥、大麥、玉米和飼料中,因此絕大部分DON檢測方法是針對這些物質,而分析穀物及飼料中的DON並非易事,許多因素必須給以良好控制。例如要從大量的原始材料中抽取適當的樣品,適當地儲備、處理以及恰當地從樣品中抽小樣。原始材料可能存在的不均一性和在儲備、分裝過程中可能出現的污染物使分析進一步複雜化;另一方面,分析過程中需要估計精確度、精密性、最低檢出限以確保檢測得出的數據能具有代表性,儘可能把分析錯誤減到最小。
