十九世紀的前生命化學思想
黑格爾
德國哲學家黑格爾(1770年-1831年)在其《自然哲學》中說,“化學過程本身是地球上的一般過程......而氣象過程則是自然界的巨大化學過程......這類過程又是生命過程的直接起源......化學過程是一種類似於生命的東西;我們在這裡遇見的生命內在活動會令人感到驚奇。化學過程如能自動地繼續進行下去,那就會成為生命;因此,顯然應該從化學方面理解生命”(黑格爾1986) 。
錢伯斯
英國業餘博物學家錢伯斯(1802年-1871年)在於1844年匿名出版的《自然創造史的遺蹟》一書中說,由於化學家已經合成了尿素和尿囊素,就再也不用證明有機化合物具有獨特的化學特性了。他認為細胞是“無機物和有機物之間的交匯點,它是礦物界的結尾和動物界的開端”,這樣,生命的創造就歸結到了“第一個原始細胞是如何產生的”這樣一個問題。他提出,某種“化學—電的作用”導致了第一個細胞的形成(瑪格納2001)。
達爾文
1871年,英國博物學家達爾文在寫給英國植物學家胡克(Joseph Dalton Hooker)的一封信中談到了生命起源的問題,他寫道,“人們常說初次產生生物的一切條件現在都具備,過去也會是如此。然而如果(好傢夥!這是多么偉大的如果!)我們能夠想出某一個溫和的小池塘,並有氨、磷酸鹽、光、熱、電等所有的東西,並想像形成了某種蛋白質似的化合物,準備經歷更複雜的變化;在現在情況下這樣的物質將會立刻被吞食或吸收,這種情形在生物形成之前是不會發生的”(邁爾1990) 。
達爾文關於生命可能起源自溫暖小水池的觀點現在都還受到一些學者的追捧。雖然20世紀的實驗科學已經證實,在合適的光、熱或電的條件下,由無機物(氨等)確實可以形成有機分子(胺基酸、肽類),但這實際上離揭示真正的生命起源還相差甚遠。
美國化學家夏皮羅(Robert H. Shapiro,1935年-2004年)在1986年的著述中注意到達爾文上面這段似乎不經意的說法,“竟然與今天的知識相當符合,如果不是因為他有遠見,就是說明多年來生命來源的研究沒有什麼進步”(詹腓力1999)。
華格納
德國探險家和博物學家華格納(Moritz Wagner,1813年–1887年)在1874年出版的《自然科學的爭論問題》第1卷中指出:在這旋轉著的星雲中,“在各不相同的區域中有各不相同的溫度,所以完全有理由假定:為有機的生命所必需的各種條件,總可以在其中的某個地方找到。因此,我們可以把天體和旋轉著的宇宙星雲的大氣看作有生命形式的永久儲藏所,看作有機胚種的永恆栽培地”(恩格斯1925)。雖然華格納給予的是一種巨觀的原則性論述,但是,即使是在今天,這種關於生命起源客觀性的見解也是毋庸置疑的。
恩格斯
德國哲學家恩格斯(1820年-1895年)在《自然辯證法》中說,“當行星有了一層硬殼而且在它的表面上有了積水的時候,行星固有的熱就比中心天體傳送給它的熱愈來愈減少。它的大氣層變成我們現在所理解的意義下的氣象現象的活動場所,它的表面成為地質變化的活動場所……最後,如果溫度降低到至少在相當大的一部分地面上不越過蛋白質能在其中生存的限度,那么在其他適當的化學的先決條件下,有生命的原生質便形成了……也許經過了多少萬年,才出現了可以進一步發展的條件,這種沒有定形的蛋白質能夠由於核和膜的形成而產生第一個細胞……生命的起源必然是通過化學的途徑實現的”(恩格斯1925) 。
恩格斯試圖從行星環境演化與生命構建物質(蛋白質)之間的關係推測生命可能的起源方式,並強調了(第一個)細胞起源的重要性。當然,那時對原核生物還沒有認識,因此,最初的細胞並不可能是如恩格斯所說的有細胞核的細胞。此外,那時也不可能知道構建細胞的生物大分子除了蛋白質外,還有核酸(DNA和RNA)。
二十世紀的前生命化學研究
奧巴林的“團聚體假說”
蘇聯生物化學家奧巴林(Alexander Ivanovich Oparin,1894年~1980年)於1936年出版了《地球上生命的起源》一書,提出了關於生命起源的化學進化理論。他將生命的起源分為三個階段:首先,原始地球大氣是還原性的,主要由CH、NH、HO、HS、CO或CO、PO等組成(O和O稀少或缺乏),短波紫外線照射能催化這些無機分子生成構成生命的一些小分子有機物(如胺基酸),接著,小分子有機物可聚合成生物大分子並在原始海洋中進一步形成多分子體系的團聚體(如胺基酸可聚合成多肽和蛋白質、一定長度的磷脂能形成構成細胞膜的脂質雙層),最後,在自然的選擇作用下,適合於當時外界條件的團聚體得以保存,不適者則消亡,經過漫長的演化,終於出現了能夠新陳代謝和自我複製的原始生命—原生體,並最終產生了細胞。
奧巴林提出了原始大氣是還原性的假設以及高能的紫外線可促進像胺基酸這樣的小分子有機化合物生成的觀點,這是一個重要的進步,也受到大量現代科學研究證據的支持。但是,在其它方面,奧巴林的觀念並不比達爾文和恩格斯前進了多少,因為,如何從大分子聚合物(奧巴林所說的‘團聚體’)進化到能夠自我複製的細胞仍然沒有答案。
米勒的實驗
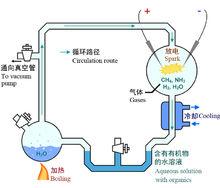 著名的米勒實驗裝置
著名的米勒實驗裝置在1952年,美國化學家米勒(Stanley Lloyd Miller,1930年-2007年)用實驗驗證了奧巴林關於在還原性大氣環境下小分子有機化合物起源的假說。米勒在一個大燒瓶中放入早期地球大氣中可能有的化學元素:水、氨、甲烷和氫,然後釋放電火花,幾天之後,在燒瓶底部聚集起一種橙色黏稠混合物,在甲烷中的碳有10-15%跑到有機化合物那裡去了,包括構成蛋白質的9種胺基酸,這樣通過向無機混合物的前生命湯放電,創造出了一些生命物質,雖然不是生命本身,但是,是一些能使生命成為可能的化合物(格萊澤2011)。無聲放電是使用臭氧發生器進行的,用氮氣代替氨進行火花放電也得到同樣的結果,可胺基酸的產量低。米勒認為醛、氨及氰化物發生反應生成氨基氰,再被水解成為胺基酸,即是按斯特雷克爾反應進行的(原田1978) 。
米勒之後的實驗
以後其他學者又進行了大量的模擬實驗,或改用緊外線、b射線、高溫、強的陽光等作能源,或改換了還原性混合氣體的個別成分(如以H2S代替HO、以HCN代替CH和H、或增加CO、CO等),結果都能產生胺基酸;而用氧化性混合氣體代替還原性混合氣體進行實驗,則不能生成胺基酸。現在組成天然蛋白質的20種胺基酸,除了精氨酸、賴氨酸和組氨酸以外,其餘的都可用模擬實驗的方法產生。組成核酸的生物小分子多數亦能通過模擬實驗形成;如有人用紫外線或g射線照射稀釋的甲醛(HCHO)溶液獲得了核糖和脫氧核糖;用紫外線照射HCN獲得了腺嘌吟和鳥嘌吟;用丙炔腈(N≡C-C≡CH)、KCN和HO在100℃下加熱一天得到了胞嘧啶;將NH、CH、HO和蘋果酸與聚磷酸加熱至100~140℃獲得尿嘧啶;將腺嘌呤及核糖的稀溶掖與磷酸或乙基偏磷酸鹽(ethylmetaphosphate)放在一起,用紫外線照射,可生成腺苷;將腺苷、乙基偏磷酸鹽封入石英玻璃管中用紫外線照射,可產生出腺苷酸(AMP)等。此外脂肪酸也可通過用高能電子照射碳氫化合物和二氧化碳而獲得。
從前生命到細胞——生命的光化學起源
沒有能量就沒有生命,而地球上支撐生命最普遍的能量來源就是太陽光。因此,最初的生命形式必定是以光的感知、傳遞、轉化和利用為核心的,這是原始細胞出現與運行的客觀目的。
從感光分子到光合色素
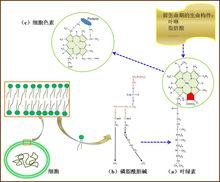 葉綠素與細胞色素的血紅素輔基
葉綠素與細胞色素的血紅素輔基可以從感光分子到光合色素來追尋早期生命的蹤跡。在尚處於生命前夜的有機湯中,應該具備了形成光合作用最重要的物質即葉綠素(或細菌葉綠素)的基礎—含有26個π電子的高度共軛的卟啉環及一個能與之連線的長度約為20個C的烷鏈(請注意,構成細胞膜的磷脂中的長鏈脂肪酸約含14-24個C,葉綠素分子的‘尾巴’或許就來自這樣的膜結構成分,雖然這只是猜想)。這種卟啉分子既能通過共振傳遞光能,又能傳遞電子。
正是生命構件特別是光合色素分子之間的偶然碰撞在渺茫的幾率中頑強而持續地進行著,才有可能形成能夠感受與捕捉最後到利用光能的一種機制。現已知道,在現代光合生物中,葉綠素的生物合成過程中的中間產物以及游離的葉綠素分子是可以吸收光能的,這些天生的“感光分子”似乎與生俱來就具有通過光來改造與衍生自我的化學潛能—這不就是最終發展成生命的那種潛能嗎?只要有光能的激發,這些分子就會反覆地產生,並在無數次偶然的相互碰撞中試探反應與利用能量的新組合,探尋通向葉綠素分子誕生的成功之旅。
當然剛開始肯定不可能會有什麼酶的催化,一切可能都是在太陽光能的驅動下的自然的物理化學作用過程。現存生物中,葉綠素的生物合成需經歷17個酶促反應步驟或更多,但需要指出的是,它卻始於一種普通而常見的胺基酸—谷氨酸,這或許具有重要的意義,因為谷氨酸是容易自然形成的常見的胺基酸之一,也是構成蛋白質的主要胺基酸之一。可以推測,可能應該是先有像類似葉綠素這樣的感光分子,然後在光合系統的不斷最佳化過程中逐漸形成了用常見的簡單組分(如谷氨酸)通過一系列酶促反應來合成葉綠素的複雜但高效的途徑。因此,真正的光合作用起源之前,像類似卟啉環這樣複雜的感光分子肯定不會稀少,但若要持續地滿足光合作用的需求,肯定還需要建立從更普遍、更簡單的原料來合成的途徑,這可能就是我們現在見到的葉綠素的合成途徑。這看上去像一種智慧型設計,但實則是一種方向性篩選的產物。
細胞被膜與光系統的協同演化
在現代生物的光合系統中,所有葉綠素分子必須和蛋白質結合才能行使光能的傳遞與利用之功能。因此,在真正意義上的光合作用出現之前,在感光分子與多肽/蛋白質之間,必定經歷了極為漫長的相互作用歷程,而且必須是在相對封閉的條件下。
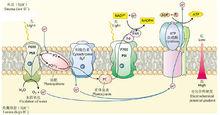 膜偶聯的光系統和電子傳遞鏈
膜偶聯的光系統和電子傳遞鏈可能首先必須有細胞被膜的前身—脂質的囊泡的出現,這可視為前細胞體。前面也談過,這樣的脂質可能提供了一種有利於通過脫水的聚合反應的微化學環境,這樣就有利於胺基酸脫水聚合成多肽鏈(甚至對核酸也是如此)。因此,當像卟啉這樣的“感光分子”連同一些多肽/蛋白被包埋於這樣的囊泡膜之中時,雖然極其偶然但如果能出現相互作用機會的話,也許就能產生出一些類似於葉綠素的色素分子(可能從質膜的長鏈脂肪酸獲取“尾巴”),而這樣的色素分子與多肽鏈之間反覆的相互作用說不定就發展成了現在的色素蛋白複合體的前體,後來逐漸演變成現在見到的定位於膜上的色素蛋白複合體。
細胞誕生之前,自然界應該具備了形成原始細胞的物質基礎。構想一下,在數十億年前的古時候,在某個海岸附近存在了一些富含有機分子(有既疏水也親水的兩親分子如脂肪酸、還有親水性的大分子化合物如多肽、以及疏水性的大分子化合物如長鏈脂肪烴等)的小水體中,偶爾一些聚集成團的兩親分子在靜靜的水面上形成無數微小的雙層膜囊泡。在無數這樣的囊泡中,總有一些會與包埋或黏附其中的感光分子和其他生物大分子(如多肽)在光能的驅動下發生物理—化學性的相互作用。這樣的過程既是偶然的,也是必然的。
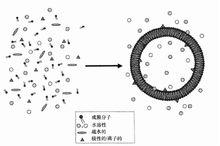 疏水分子自發裝配成囊泡
疏水分子自發裝配成囊泡或許在無數次這等原始囊泡的形成過程中,在囊泡內部偶爾含有過一些能夠利用太陽光將無機元素合成簡單有機化合物的複雜有機分子或分子聚集體,終於發展成能夠進行光合作用的機構。譬如,在前細胞的膜上,或許色素分子與多肽之間反覆的相互作用就會向色素蛋白複合體方向不斷完善(更有效地接受和傳遞光能),或者說,在無數次隨機組合事件中,偶爾錨定於膜上的一些色素—多肽複合體由於能較好地吸收與傳遞光能,就被選擇下來了並進一步發展成光反應中心。可能最先出現的是像PSI這樣稍微簡單一些的結構,而複雜的裂解水功能可能是之後獲得的。偶爾有些蛋白能利用膜內外的H梯度將ADP轉化成ATP,於是就發展成了ATP酶,這種酶在放氧複合體出現後可能得到了更大的青睞。另一方面,一些核苷酸由於能攜帶胺基酸偶爾有助於那些色素蛋白複合體中的多肽的形成也受到了選擇,或者說,一些類核酸的組合體偶爾獲得了合成肽鏈的能力,於是就被選擇下來並得到不斷完善。
在原始海洋中作為原始生命前體的大分子化合物的形成更應該是在容易接近乾濕交替狀態的沿岸帶的小型水體之中,而不大會在無邊無際的汪洋大海之中,這種過程似乎可能是:有機湯→脂質體(油滴)與感光分子—多肽—核(苷)酸等的混合物→類細胞體→細胞。此外,脂雙層結構有助於生物大分子(如RNA)的脫水縮合作用這樣的一種特性也為在相對溫和的條件下細胞的起源與進化提供了一種可能途徑。
從前生命化學到生物化學(酶的誕生)
光合作用就是通過一系列氧化還原反應將被色素分子吸收和傳遞過來的太陽光能轉化為化學能的過程。在光合作用的演化過程中,也許一些多肽或蛋白(在某些條件下)的存在,偶然加速了某些反應的進程(而自身並不參與化學反應),這種反覆的相互作用和選擇過程或許終於導致了具有催化作用的蛋白質—酶的出現。而且,一旦一種蛋白偶爾起到了催化作用,它可能就會受惠於穩定化和最佳化的選擇,直到一種具有高效催化作用的酶的誕生。
構成最簡單光合機構的基本組分首先必須存在,這是在無數組合中篩選出功能性組合的前提,只有在這樣的功能單位出現之後,沿著一種功能性目標(即最有效的光合作用)的自然選擇才可能發生,系統才會進入結構與功能的不斷最佳化過程。
一般來說,即使是放能反應也不一定會自發地進行,因為反應物首先需要被活化,即需要活化能。這是使一個化學反應中所有分子達到反應狀態所需要的能量。而催化劑就是指能夠降低一個反應活化能的物質。催化劑有一個特性,即它能夠加快反應,但自身並不發生變化。
只有一個高效的光合作用系統才能生產更多的有機物,這種光合系統就會被更廣泛地傳播或擴展—這可能就是光合系統不斷向高效方向演化的核心動力,這也是自然選擇的方向性。因此,最初的酶可能就是在與光合作用過程相關的各種氧化還原反應與多肽/蛋白質的相互作用過程中逐漸形成的,如果通過酶促反應可以不斷地提高反應速率,就可能獲得競爭與被選擇下來的優勢,這應該就是酶促反應速率能夠達到驚人速度的根本緣由。這樣,蛋白質一方面成為光合系統的結構支撐部分,另一方面成為推動一系列生化反應高效進行的催化劑,因此就出現了酶蛋白與結構蛋白之間在功能上的分化。
酶之所以受到生命的青睞,因為酶可以進化出驚人的催化能力:譬如,無機鐵將過氧化氫分解為氧氣和水時,催化效率很低,當鐵與原卟啉結合形成原血紅素時,其活性增加了1000倍,若原血紅素與過氧化氫蛋白結合,則活性提高100萬倍。酶就是生命舞台上的魔術師,它以驚人的速度,製造出令人眼花繚亂的各種各樣的複雜有機物!酶是有機物中一種主要的專業工具機。催化作用的出現,以自身作為反應物或催化其他反應,將可利用的前體轉化成產物,豐富了原始世界的化學多樣性,最終形成自動催化循環。酶的高效性一方面取決於酶的循環催化特性,即酶能被生化反應循環利用,因為催化劑與產物是分離的,所以不會被消耗,也能持續地發生作用;另一方面,酶具有高度的特異性,因為,特化是高效之前提。
當然,催化劑並不是有機物質的專利,因為很多無機物也能催化另外的無機反應,其實,很多酶還是離不開無機金屬離子的幫助。在生命之前的原始有機大分子的產生可能依賴於高能(在高溫或強輻射條件下)以及近於無水(或通過脂質創造的微化學環境)條件下的脫水反應,而生命的進化則產生出了有機大分子脫水劑(酶),依賴它成功獲得了在溫和的稀溶液中進行脫水反應的能力。這不能不說是生命進化的偉大創舉。但是,實在難以想像,如果沒有生命選擇的方向性以及個體化的生命環境(如細胞隔膜),這種酶的進化如何得以實現?如何駕馭數以千計的蛋白酶類而建立起一個有序的生命體系?
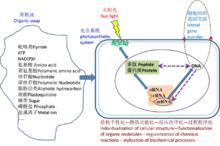 細胞的化學起源
細胞的化學起源蛋白酶最大的神奇來自它通過一種非共價、低鍵能的結合力量(就像要分離粘合在一起的兩個物體,一個人從一個方向用力無法撼動,但2個以上的人從不同的方向一起用力,就容易將它撕開了)輕易地就撕開了堅固的共價鍵(如光合作用中的水分子的裂解),再通過高能化合物將有機分子活化輕易地就形成縮合的共價結合,並演化出了令人無法想像的快速反應能力,而且本身還不被消耗,可以循環利用。要知道,共價鍵的鍵能要比非共價鍵高5-10倍!正是這種神奇的化學作用方式,使在溫和環境中利用簡單物質製造複雜有機物成為可能(稱之為光合作用),從而為細胞的起源和演化之旅得以開啟奠定了生化反應的基礎 。
生物化學的核心——遺傳系統的成型
前生命化學的最終階段就是將遺傳物質指揮生命構建過程程式化,即將生命構建過程信息化地儲存於DNA之中,這是個性化的生命獲得自我繁殖特性必須邁出的重要一步 。最後是細胞分裂機制的逐漸形成,這得益於細胞膜的半通透性(小分子養分的進入與大分子碳水化合物的堆積),以及二者之間的矛盾及其必然的結果—細胞破裂過程的無數次重複。精確而周期化的細胞分裂才能使穩定的遺傳成為了可能,這時像我們在現存生物中能所見到的細胞才成功地誕生了,細胞運行機制才得以程式化了。
