
主要凝血因子
因子 I, 纖維蛋白原
因子 II,凝血酶原
因子 III,組織因子
因子 IV,鈣因子(Ca2+)
因子 V,促凝血球蛋白原,易變因子
因子 VII,轉變加速因子前體,促凝血酶原激酶原,輔助促凝血酶原激酶
因子 VIII,抗血友病球蛋白A (AHG A),抗血友病因子A (AHFA),血小板輔助因子 I,血友病因子 VIII 或 A,
因子 IX,抗血友病球蛋白B (AHG B),抗血友病因子B (AHF B),血友病因子 IX 或 B
因子 X,STUART(-PROWER)-F,自體凝血酶原C
因子 XI,ROSENTHAL因子,抗血友病球蛋白C
因子 XII,HAGEMAN因子, 表面因子
因子 XIII,血纖維穩定因子
輔助凝血因子
共同凝血
FITZGERALD因子
FLETCHER因子 (激肽釋放酶原)
von-Willebrand-因子
被取消資格的凝血因子
因子 VI,促凝血球蛋白:其實是活化後的第五因子。
這些因子共同作用,會導致凝血。
如果一種或多種凝血因子缺失,會導致“血友病”: 血液病,血友病。
列舉
同義語 縮寫符號 血漿濃度 在血清中 貯存穩定性 參與凝血途徑
Ⅰ 纖維蛋白原 Fg 2000-4000 無 穩定 共同
Ⅱ 凝血酶原 200 有10%-12% 穩定 共同
Ⅲ 組織凝血激酶TF 外源
Ⅴ 前加速素 5-10 無 不穩定 共同
Ⅶ 前轉變素 2 有 穩定 外源
Ⅷ 抗血友病因子 AHG 〈10 無 不穩定 內源
Ⅸ 血漿凝血激酶PTC 3-4 有 較穩定 內源
Ⅹ Stuart Power因子 6-8 有 穩定 共同
Ⅺ 血漿凝血激酶前質 PTA 4 有 穩定 內源
Ⅻ 接解因子 HF 2.9 有 穩定 內源
PK 激肽釋放酶原 PK 1.5-5.0 有 穩定 內源
HMWK 高分子量激肽原 HMWK 7 有 穩定 內源
ⅩⅢ 纖維蛋白穩定因子 FSF 25 無 穩定 共同
凝血因子Ⅷ的結構以及性質
在血友病中,80%的病人患有血友病A,該類病人主要缺少正常人所擁有的凝血因子Ⅷ。
FⅧ基因位於X 染色體長臂末端(Xq28), 長為186kb,由26個外顯子組成,其中第l4號外顯子為3.1 kb,是目前發現的人類最大的外顯子之一.FⅧ mRNA長9kb,編碼一條由2351胺基酸組成的前體多肽.去除N端l9個殘基的個信號肽後,成熟蛋自由2332胺基酸組成.胺基酸順序分析表明,FⅧ 蛋白由3個A 結構域,一個B結構域及2個C結構域組成.各區的排列順序為A1-A2-B-A3-C1-C2 ,從血漿中及從重組細胞培養上清中分離純化得到的FⅧ 是由兩條肽鏈所組成, 重鏈由A1-A2-B或A1-A2組成, 分子量為90~200kDa 不等; 輕鏈由A3-C1-C2所組成,分子量為80kDa,二條肽鏈間由Ca2+連線。目前的研究表明,B結構域對FⅧ的凝血活性並非必需。
正常人血漿中的Ⅷ因子是一種分子量高達100萬~200萬的糖蛋白,含低分子量及高分子量兩種成分。低分子量具有凝血活性(Ⅷ∶C),而高分子量成分具有Ⅷ因子的相關抗原(ⅧR:Ag)及VW因子(ⅧR∶VWF),這三種成分的合成部位,控制其合成的基因位點及遺傳方式均不相同,Ⅷ∶C由肝脾或單核細胞所合成,受X染色體遺傳控制,在血友病甲中Ⅷ∶C活性降低,為性染色體隱性遺傳。ⅧR∶Ag及ⅧR∶VWF由內皮細胞、巨核細胞及血小板合成,由常染色體遺傳。正常血漿VWF值為10mg/L,其活性由一系列血漿多聚體產生,多聚體的分子量約40萬~2000萬以上,存在於血漿,血小板內及血管內皮下,血漿VWF濃度輕度下降或高分子量多聚體選擇性喪失,可使血小板粘附功能降低。在VWD患者中缺乏這兩種成分,或表現為這兩種成分的分子結構異常,Ⅷ因子高分子的各種生理活性異常,Ⅷ∶C功能也有相應的影響。ⅧR∶Ag中含有低、中、高分子量不同的聚合物,構成VWD的不同亞型。ⅧR∶Ag的高分子聚合物通過與血小板的特殊受體結合可使血小板粘附於血管內皮下層,維持正常的出血時間,瑞斯托黴素和ⅧR∶VWF對血小板的作用是與血小板的受體有關。本病患者的Ⅷ∶C和ⅧR∶Ag,也有不同程度降低,當輸入因子Ⅷ提純製劑後,患者的出血時間在短時有所縮短,而Ⅷ∶C則6~24小時後才上升。目前認為正常的ⅧR∶Ag有穩定Ⅷ∶C的作用,當ⅧR∶Ag缺乏時即可影響Ⅷ∶C的活性,故本病可能是由於Ⅷ因子的高分子部分有缺陷,或是整個因子Ⅷ複合物的缺陷所引起。最近發現VWD的纖維蛋白溶解作用也有缺陷。
凝血因子Ⅷ的測定
根據中華人民共和國藥典三部 附錄X N 人凝血因子VIII測定法(一期法)
本法系用人凝血因子VIII缺乏血漿為基質血漿,採用一期法測定供試品人凝血因子VIII效價。
試劑:(1)3.8%枸櫞酸鈉9.5g,加水溶解並稀釋至250ml。
(2)咪唑緩衝液(pH7.3)取咪唑0.68g和氯化鈉1.17g,加水使溶解成100ml,加入0.1mol/L鹽酸溶液42.2ml,再加水稀釋至200ml,即得。
(3)稀釋液 取一體積的3.8%的枸櫞酸鈉加入5體積咪唑緩衝液混合,加適量20%人血白蛋白至終濃度為1%
(4)激活的部分凝血活酶(APTT)試劑
(5)人凝血因子VIII缺乏血漿 為人凝血因子VIII含量低於1%的人血漿或人工基質血漿
(6)0.05mol/L氯化鈣溶液 取氯化鈣(CaCl2?2H2O)147g,加水溶解並稀釋至1000ml,配製成1mol/L的氯化鈣貯存液,用前用水稀釋20倍,配製成0.05mol/L氯化鈣溶液。
人凝血因子VIII標準液的製備 用人凝血因子VIII缺乏血漿將標準品稀釋成每1ml含1IU凝血因子VIII,再用稀釋液分別進行10倍、20倍、40倍和80倍稀釋,置冰浴待用。
供試品溶液的製備 用人凝血因子Ⅷ 缺乏血漿將標準品稀釋成每1ml含1IU凝血因子Ⅷ,再用稀釋液進行10倍和20倍或40倍稀釋,置冰浴待用。
測定法 取激活的部分凝血活酶試劑0.1ml,置37℃水浴保溫一定時間(一般4min),加人凝血因子Ⅷ缺乏血漿0.1ml、供試品溶液0.1ml,混勻,置37℃水浴保溫一定時間(一般5min),加已預熱至37℃ 0.05mol/L氯化鈣溶液0.1ml,記錄凝固時間。
用不同稀釋度的人凝血因子Ⅷ標準溶液0.1ml替代供試品溶液,同法操作。
將標準溶液人凝血因子Ⅷ效價(IU/ml)的對數對其相應的凝固時間(秒)的對數進行直線回歸處理,求出直線回歸方程。計算供試品溶液人凝血因子Ⅷ效價,再乘以稀釋倍數,即為供試品人凝血因子Ⅷ效價(IU/ml)。
臨床套用
PT延長
通常認為PT延長代表凝血因子Ⅱ、Ⅴ、Ⅶ、Ⅹ的活性低於正常或抗凝物質的存在。肝功能輕度受損,PT仍可正常,它僅在肝實質細胞嚴重損害時才明顯延長。僅以PT判斷肝病患者凝血功能異常和肝細胞損傷程度是不夠的,如同時測定凝血因子的活性,可能更有價值。
肝病與凝血因子Ⅱ
大多數研究認為急性肝炎和慢性肝炎輕度患者,凝血因子Ⅱ活性正常或輕度下降;慢性肝炎中度、重度和肝硬化患者,凝血因子Ⅱ活性水平明顯下降,說明它的降低程度與肝細胞損害程度密切相關。有研究認為異常凝血酶原(protein-Ⅱinduced by vitamin Kabsence,PIVKA-Ⅱ)可用於原發性肝癌的診斷,部分AFP陰
性的原發性肝癌患者PIVKA-Ⅱ陽性,還有研究認為小肝癌患者PIVKA-Ⅱ陽性率高於AFP,它還有助於原發性肝癌的病情變化及療效判斷,在臨床上應聯合檢測AFP與PIVKA-Ⅱ。
凝血因子Ⅴ
 凝血因子
凝血因子研究顯示凝血因子Ⅴ活性在肝功能失代償或嚴重肝病時才減少,故認為它是判
斷肝病患者預後的良好指標。Izumi等研究顯示:對乙醯氨基酚誘導的需肝移植的暴發性肝功能衰竭患者,凝血因子Ⅴ活性<20%時對死亡的陽性預測值為0.49,
<10%時為0.57;而其它原因誘導的需肝移植的暴發性肝功能衰竭患者,凝血因子Ⅴ活性<20%時對死亡的陽性預測值為0.85,<10%時為1.00,因此認為凝血因子Ⅴ活性是判斷非對乙醯氨基酚誘導的暴發性肝功能衰竭患者預後的最佳預測指標。鄒正升等研究認為凝血因子Ⅴ水平比PTA更特異的反映重型肝炎患者的預後,兩者聯合可能有助於更早更準確診斷重型肝炎,同時指出應加強重型肝炎因子Ⅴ的檢測及重視因子Ⅴ在作為肝衰竭患者行肝移植術時的主要篩選指標的研究。凝血因子Ⅴ活性除用於判斷預後外,還與血栓的形成密切相關,可作為門靜脈血栓形成的預測指標。
凝血因子Ⅶ
凝血因子Ⅶ的半衰期最短(4~6h),血漿含量較低(0.5~2mg/L),故可作為肝病患者蛋白質合成功能減退的早期診斷指標。Rodriguez-Inigo等在慢性肝病患者通過肝活檢組織原位雜交的方法檢測到凝血因子Ⅶ的表達與肝纖維化的分級呈負相關,可作為預測纖維化程度的指標。凝血因子Ⅶ活性還與預後有著密切的聯繫,如Violi等研究認為凝血因子Ⅶ活性<34%的肝硬化患者93%在隨訪10月內死亡,故認為它是肝硬化患者預後好壞的早期預測指標,可更好識別肝移植候選人。肝硬化患者凝血因子Ⅶ活性可明顯下降,凝血因子Ⅶ缺乏可導致血小板活性的改變,結合血小板計數減少使出血時間延長,因此對有創診斷與治療的肝硬化患者,還應該用凝血因子Ⅶ活性進行出血危險度的評估,而不能僅看血小板計數。除診斷之外,重組凝血因子Ⅶ可以有效地糾正肝病患者凝血異常,有利於有創性檢查的進行。
凝血因子Ⅷ
凝血因子Ⅷ不僅由肝細胞產生,而且由竇內皮細胞與庫普弗細胞產生,其它組織如腎臟也可產生。當肝細胞合成功能減退時,竇內皮細胞及庫普弗細胞仍維持凝血因子Ⅷ的合成;肝臟清除功能減退,內毒素及免疫因素刺激使它的合成與釋放增加。范威氏因子(vonwillebrandfactor,vWF)主要由肝外合成,肝硬化患者可能由於內毒素血症,血管內皮細胞功能異常,使其釋放增加;vWF分解蛋白酶對其分解減少,也使其血漿水平升高。在大多數病毒性肝炎患者凝血因子Ⅷ活性、vWF均明顯升高。但肝病合併DIC者,由於凝血因子大量消耗,使凝血因子Ⅷ活性水平降低,故中國將凝血因子Ⅷ活性小於正常50%作為診斷肝病合併DIC的必備條件之一。
參與血液凝固過程的各種組分;其中大多是含糖的絲氨酸蛋白酶。整個凝血過程大致上可分為兩個階段,凝血酶原的激活及凝膠狀纖維蛋白的形成。
凝血酶原激活
簡介
 激活
激活體記憶體在有內源性及外源性兩種激活系統。前者是指心血管內膜受損,或血液流出體外通過與異常表面接觸而激活因子Ⅻ(Hageman factor)。後者則由於組織損傷釋放出因子Ⅲ,從而激活因子Ⅶ。兩者都能啟動一系列連鎖反應,並在因子Ⅹ處匯合,最後都導致凝血酶原的激活及纖維蛋白的形成。
內源性激活系統
整個凝血酶的激活途徑如圖2所示。當血液與帶負電荷的膠原蛋白(皮膚血管外壁)或異體表面(如高嶺土、玻璃等)接觸時,因子Ⅻ就由酶原激活成Ⅻa,後者除能激括因子Ⅺ外,又同時使血漿前舒緩激肽釋放酶激活。激活後的激肽釋放酶在高分子量激肽原的促進下反過來又進一步使因子Ⅻ激活,但此時不再是接觸激活而是肽鍵水解激活(見蛋白水解酶),使成為因子Ⅻf。這是一正反饋效應,不論Ⅻa或Ⅻf都具有相同的活力。激活後的Ⅻa在Ca2+存在下接著又使因子Ⅸ激活。因子Ⅻ是由596個胺基酸殘基所組成,因子Ⅺ是由兩個亞基所組成,每一亞基含607個胺基酸殘基,其結構與血漿激肽釋放酶很類似。
因子Ⅸ由416個胺基酸殘基所組成,激活時釋放出一肽段,形成由二硫鍵連結的兩條肽鏈。與磷脂結合的部位在輕鏈,而酶的催化活性部位則在重鏈。活化的因子Ⅸa在Ca2+與磷脂存在下與因子Ⅷ形成複合物,使因子Ⅹ激活為因子Ⅹa。在正常生理條件下磷脂由血小板提供,在此反應中因子Ⅸa起酶催化作用,而因子Ⅷ只是起調節作用,由於它也能與因子Ⅹ結合,從而使局部的底物濃度增高。事實上單獨因子Ⅸa也能使因子Ⅹ激活,但在因子Ⅷ參與下反應速度可增加數千倍以上。因子Ⅷ還需有因子Ⅹa及凝血酶的激活而成為因子Ⅷ',這裡也是一正反饋效應。因子Ⅷ是一分子量達百萬以上的糖蛋白,高鹽濃度下解離成分子量約20萬的亞基。若體內由於基因缺陷,因子Ⅷ欠缺或無活性,在臨床上就表現出先天性血友病。因此因子Ⅷ又稱為抗血友病因子。
因子Ⅹ是由448個胺基酸殘基所組成,激活時釋放出一肽段,形成由二硫鍵連結的兩條肽鏈。它與因子Ⅸ相似,與磷脂及因子Ⅴ的結合部位在輕鏈,而酶的催化活性部位在重鏈。
激活後的因子Ⅹ與Ca2+、磷脂及因子Ⅴ共同形成一複合物,後者最終使凝血酶原激活為凝血酶。因子Ⅴ的性質與因子Ⅷ有很多相似之處,它不是起酶的催化作用,而是加速凝血酶原的激活,當因子Ⅴ與磷脂同時存在時激活過程可加速2萬倍。同樣因子Ⅴ也可被凝血酶激活成Ⅴ',成為另一正反饋效應。因子Ⅴ也是一大分子量的糖蛋白,由分子量約30萬的亞基所組成,在體內極不穩定,容易被體內蛋白C(也是一種絲氨酸蛋白酶)所破壞,因此稱為不穩定因子。
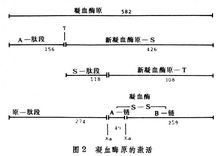
凝血酶原(即因子Ⅱ)由 581個胺基酸殘基所組成,當被因子Xa複合物激活時,幾乎同時在肽鍵Arg(精274)-Thr(蘇275)及Arg(精322)-Ile(異亮323)處水解,並自N端釋放出分子量約3萬的肽段(殘基1~274),形成由兩條肽鏈通過二硫鍵連線的凝血酶。激活後的凝血酶又能催化降解凝血酶原,在殘基Arg(156)-Ser(157)處的肽鍵水解,釋放出A肽段並形成新凝血酶原-S,後者就不易再被Xa所激活。有人認為片段A通過Ca2+及磷脂與因子Xa相結合,如果此肽段被水解除去後,新凝血酶原-S就喪失與因子Xa結合的能力,即使它仍含有可被因子Χa專一水解的肽鍵,反應也極不易進行。這是凝血酶原激活過程的一個重要的負反饋調節機制,避免了體內由於產生過量凝血酶而引起血栓。
當凝血酶原激活時從N端釋放的肽段,大致上可分為兩個區域,即A肽段(殘基1~156)及S-肽段(殘基157~274)。此兩肽段在胺基酸組成上特別是二硫鍵的位置非常相似,其中有31個胺基酸殘基完全相同,在構型上似乎各自成為獨立的單位,被稱為“環餅”結構。一般認為此兩環形結構能分別與因子Xa相結合,因而可在兩個肽鍵處(殘基274~275,322~323)同時水解而激活成凝血酶。如果只有殘基 274處的肽鍵被因子Xa水解,生成的新凝血酶原-T則不能再激活成凝血酶。
外源性激活系統
 凝血因子
凝血因子體內組織損傷時釋放出因子Ⅲ,也稱為組織因子。在Ca2+存在下它能與血液中已活化的因子Ⅶ形成複合物,就能使因子Ⅹ激活,此後就與內源性激活途徑的反應步驟相同。通過外源性途徑血液凝固在10多秒鐘內即可完成,而通過內源性途徑則需數分鐘。
因子Ⅲ為一膜糖蛋白,由263個胺基酸殘基所組成,存在於血管內皮細胞,分布於體內各組織,在肺、腦、胎盤中更豐富。如果細胞膜受到損失它就隨之釋放。因子Ⅲ的作用類似於因子Ⅷ及Ⅴ,也是調節因子,不同的是,由於存在於膜上,因而無需血小板的磷脂參與。因子Ⅶ激活因子Ⅹ的機理類似於因子Ⅸ。上述內源或外源系統中各凝血因子Ⅻ、Ⅺ、Ⅸ、Ⅹ、Ⅶ,凝血酶以及與凝血系統有關的激肽釋放酶、蛋白C都屬於絲氨酸蛋白酶,它們活性部位附近的胺基酸排列順序都與胰蛋白酶極為相似,不同的是它們都是糖蛋白。
因子Ⅶ是由406個胺基酸殘基所組成,激活時不釋放出肽段,其結構與因子Ⅸ、Ⅹ很相似。
凝血因子與維生素K在凝血酶原近N端的肽段中有一種特殊胺基酸,即γ-羧基谷氨酸。由於在同一谷氨酸側鏈中含有兩個羧基,與Ca2+的親合力就特彆強。這樣,凝血酶原就可通過Ca2+再與磷脂結合,這是因子Ⅹ激活凝血酶原所必需的。若動物給以維生素K的拮抗劑,如雙羥香豆素,則在凝血酶原分子中原有的γ-羧基谷氨酸殘基又被正常谷氨酸所取代,同時凝血機能也受到損害,由此認為維生素K是作為γ羧化酶的輔酶,在凝血酶原分子中總共有10個γ-羧基谷氨酸,它們都集中於N端32個胺基酸殘基的肽段上(圖3)。這就不難理解,一旦A肽段被凝血酶自身降解而除去後,留下的新凝血酶S就很難再被因子Ⅹa所激活。除凝血酶原外其他與維生素K有關的凝血因子還有因子 Ⅸ、Ⅹ及Ⅶ,還包括使因子Ⅴ、Ⅷ失活的蛋白C。它們在N端肽段附近都有類似的順序,γ-羧基谷氨酸的位置也都不變,激活後都形成兩條鏈,輕鏈為N端部分,含有γ-羧基谷氨酸,因而能與Ca2+及磷脂相結合。重鏈為C端部分,含有酶的催化中心。
纖維蛋白的形成
從纖維蛋白原轉變為纖維蛋白大致上可分為三個階段:
纖維蛋白單體的形成
 凝血因子
凝血因子纖維蛋白原(因子Ⅰ)為一分子量約34萬的糖蛋白,
是由兩個完全相同的亞基所組成,每一亞基又含有三條肽鏈,即α、β、γ 鏈,彼此通過二硫鍵相互連線。此三條肽鏈分別含610、461及410個胺基酸殘基。兩個亞基在肽鏈N端附近再通過三對二硫鍵將對稱的二亞基連結起來(圖4)。因此整個纖維蛋白原分子可用 (Aα,Bβ,γ)2來表示,A、B分別代表被凝血酶自α、β肽鏈N末端水解釋放的肽段,形成纖維蛋白後則用(α、β、γ)2來表示。在纖維蛋白分子中二硫鍵的位置相當集中,存在有所謂“二硫鍵節”的結構,其位置也靠近肽鏈的N端(圖4)。β與γ肽鏈的胺基酸順序很相似,特別近C端附近約有1/3是相同的。有人根據纖維蛋白原的理化性質提出了圖5 的模型,球體間的連線部分即為螺鏇區,由三條肽鏈形成繩索狀的螺鏇結構。肽鏈C端球體的大小與形狀類似血漿白蛋白,結構較緊密,並連線一條鬆散的α鏈C端肽段,容易被纖溶酶或其他蛋白酶所降解。
當凝血酶作用於纖維蛋白原時首先自 α鏈的 N端處釋放出一16肽的肽段A,經過一滯後期後自β鏈的N端開始加速釋放出一14肽的肽段B,剩下的部分即為纖維蛋白的單體。不同種屬的纖維蛋白原A、B肽段的水解位置都在Arg(精)-Gly(甘)肽鍵上。肽段A、B的胺基酸組成可因不同種屬而有很大差異,但都帶有2~6個負電荷,並含有某些特殊胺基酸,如肽段A中含有帶磷酸基的絲氨酸,肽段B中含有帶硫酸基的酪氨酸。正因為肽段A、B帶有淨負電荷,使纖維蛋白原分子在未經凝血酶降解前,由於靜電相斥而不能聚合。
纖維蛋白單體的聚合
在纖維蛋白單體的聚合過程中肽段A的釋放起主要作用,先是首尾聚合,而肽段B的釋放能使聚合加速並開始側向聚合。纖維蛋白單體由於A、B肽段的釋放,在每一亞基中暴露出兩個相嵌的互補區,單體間就可借非共價鍵首尾或側向聚合,隨著側向聚合程度加深,血塊顯得粘稠,由透明轉向不透明。
纖維蛋白的交聯
 凝血因子
凝血因子激活後的凝血酶除降解纖維蛋白原釋放肽段A、B外,
在Ca2+存在下又同時迅速使因子ⅩⅢ激活,後者能使聚合的纖維蛋白在鄰近的肽鏈間形成橋鍵,而成為穩定而交聯的纖維蛋白多聚體,即使在5M尿素溶液中也不溶解,而交聯前的凝膠在此條件下則可溶解。因子ⅩⅢ為轉谷氨醯胺酶,它使肽鏈間賴氨酸殘基上的ε氨基與谷氨醯胺殘基上的γ醯胺基連結成新的肽鍵。每一纖維蛋白單體最多能形成6個共價橋鍵,若每分子內有2~3個,就可形成很穩定的交聯纖維蛋白。
長期的進化使纖維蛋白原成為理想的止血劑,如在未激活前分子間由於靜電相斥不能聚合而成為溶膠;位於肽鏈N端的A、B肽容易被凝血酶水解除去,隨之靜電效應消失,凝膠迅速形成;在肽鏈C末端附近又可再形成橋鍵,使成為穩定的凝膠並有足夠的機械強度;分子量大,親水性強、呈對稱性,符合凝膠特性;分子中含一段繩索狀螺鏇結構區,容易被蛋白酶降解,在體內不致於形成血栓;纖維蛋白凝膠的降解產物具有抑制凝血酶的活力,也能阻止纖維蛋白單體的聚合,從而起到自身調節的反饋作用。
在凝血體系中除了各因子間的正負反饋及自身調節外,屬於蛋白酶的凝血因子又受血漿中相應的蛋白酶抑制劑的制約,例如血漿中的抗凝血酶Ⅲ(ATⅢ,antithro-mbinⅢ),除能專一抑制凝血酶外,還能抑制因子Ⅹa、Ⅸa、及Ⅶa,特別對Ⅹa的抑制效果尤其顯著。肝素能大大加速AT-Ⅲ的抑制作用,因而在臨床上被用作重要的抗凝劑。除AT-Ⅲ外血漿中還有其他蛋白酶抑制劑,如α1抗蛋白酶、抗纖溶酶及α2巨球蛋白等,它們對凝血因子中的各蛋白酶也都有一定程度的抑制作用。
相關疾病
血友病(Hemophilia)是一組由於血液中某些凝血因子的缺乏而導致患者產生嚴重凝血障礙的遺傳性出血性疾病,男女均可發病,但絕大部分患者為男性。包括血友病A(甲)、血友病B(乙)和因子XI缺乏症(曾稱血友病丙)。前兩者為性連鎖隱性遺傳,後者為常染色體不完全隱性遺傳。血友病在先天性出血性疾病中最為常見,出血是該病的主要臨床表現。治療方法包括局部止血、替代療法等。後者中新興的療法包括重組人凝血因子(注射用重組人凝血因子VIIIa,諾其)治療,因其無人工污染,安全性高的特點,有日後普及的趨勢。
