
簡介
 micro RNA
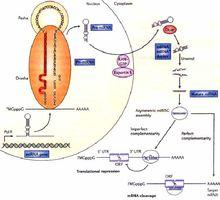
micro RNAMicroRNA (miRNA) 是一類內生的、長度約為20-24個核苷酸的小RNA,其在細胞內具有多種重要的調節作用。每個miRNA可以有多個靶基因,而幾個miRNA也可以調節同一個基因。這種複雜的調節網路既可以通過一個miRNA來調控多個基因的表達,也可以通過幾個miRNA的組合來精細調控某個基因的表達。據推測,miRNA調節著人類三分之一的基因。最近的研究表明大約70 %的哺乳動物miRNA 是位於TUs區( transcriptionunits , TUs ) ( Rodriguez et al ,2004) , 且其中大部分是位於內含子區( Kim &Nam , 2006) 。一些內含子miRNA 的位置在不同的物種中是高度保守的。miRNA 不僅在基因位置上保守, 序列上也呈現出高度的同源性(Pasquinelli etal , 2000 ; Ruvkun et al , 2001 ; Lee & Ambros ,2001) 。miRNA 高度的保守性與其功能的重要性有著密切的關係。miRNA 與其靶基因的進化有著密切的聯繫, 研究其進化歷史有助於進一步了解其作用機制和功能。
MicroRNA
MicroRNA(miRNA)是一類內生的、長度約20-24個核苷酸的小RNA,幾個miRNAs也可以調節同一個基因。可以通過幾個miRNAs的組合來精細調控某個基因的表達。據推測,miRNA調節著人類三分之一的基因。
MicroRNA存在多種形式,最原始的是pri-miRNA,長度大約為300~1000個鹼基;pri-miRNA經過一次加工後,成為pre-miRNA即microRNA前體,長度大約為70~90個鹼基;pre-miRNA再經過Dicer酶酶切後,成為長約20~24nt的成熟miRNA。
實際研究中,pre-miRNA套用最早,也最廣泛,很多商業化的MicroRNA庫都是pre-miRNA形式的。近幾年來,研究發現microRNA的雙臂對成熟miRNA的形成有著十分重要的作用,所以天然的pri-miRNA形式越來越多地被研究者採用。
MicroRNAs (miRNAs)是一種大小約21—23個鹼基的單鏈小分子RNA,是由具有髮夾結構的約70-90個鹼基大小的單鏈RNA前體經過Dicer酶加工後生成,不同於siRNA(雙鏈)但是和siRNA密切相關。據推測,這些非編碼小分子RNA(miRNAs)參與調控基因表達,但其機制區別於siRNA介導的mRNA降解。第一個被確認的miRNA是線上蟲中首次發現的lin-4 和let-7,隨後多個研究小組在包括人類、果蠅、植物等多種生物物種中鑑別出數百個miRNAs。
特徵
已經被鑑定的miRNAs據推測大都是由具有髮夾結構,約70個鹼基大小形成髮夾結構的單鏈RNA前體經過Dicer酶加工後生成的,有5’端磷酸基和3’羥基,大小約21—25nt的小分子RNA片斷,定位於RNA前體的3’端或者5’端。
3個研究小組分別從線蟲、果蠅和Hela細胞中鑑定的100個新miRNAs中,有15%跨越線蟲、果蠅和哺乳動物基因組具有高度的保守性(只有有1—2個鹼基的區別)。Lau 和Bartel 實驗室的同事更加認為:所有的miRNAs可能在其他物種中具有直向同源物(Ortholog,指那些起源於同一祖先,在不同生物體中行使同一功能的基因群就可比作為一個門類,這些類似的基因被稱為“直向同源物”)。
 micro RNA
micro RNABantam 最早被認為是果蠅中參與細胞增殖的一個基因位點。已知幾個包含增強子的轉座子插入跨越這個位點的一段12.3kb區域會導致果蠅的眼和翅重複生長,而由轉座子介導的一段跨越該位點的23kb片斷缺失則導致突變果蠅個體小於野生型果蠅。Cohen和同事用一段3.85kb的片斷導入21kb片斷缺失的果蠅中使其恢復原來的大小。但是奇怪的是表達這個3.85kb片斷中的EST卻沒有同樣的效果。Cohen將這個片斷和瘧蚊Anopheles gambiae的同源序列進行比較,發現一段90bp的高度保守區,經過RNA folding program (mfold)發現這個保守序列可以形成髮夾結構,使得這個區段很象是一個miRNA的前體。這個結果經過Northern blot證實突變果蠅的幼體缺少一個21bp的bantam miRNA ,用這個90bp的mRNA前體經過一系列的“功能缺失”—“功能恢復”實驗,證實 bantam miRNA在細胞增殖中的作用。研究人員用電腦程式檢索在hid mRNA的3’非編碼區找到了bantam的3個潛在的結合位點( hid是果蠅中一個誘導凋亡的基因),並證實 bantam miRNA抑制hid 的翻譯而非轉錄。
miRNAs的表達方式各不相同。部分線蟲和果蠅的miRNA在各個發育階段的全部細胞中都有表達,而其他的miRNA則依據某種更為嚴謹的位相和時相的表達模式(a more restricted spatial and temporal expression pattern)——在不同組織、不同發育階段中miRNA的水平有顯著差異。
功能
科學家開始認識到這些普遍存在的小分子在真核基因表達調控中有著廣泛的作用。線上蟲,果蠅,小鼠和人等物種中已經發現的數百個miRNAs中的多數具有和其他參與調控基因表達的分子一樣的特徵——在不同組織、不同發育階段中miRNA的水平有顯著差異,這種miRNAs表達模式具有分化的位相性和時序性( differential spatial and temporal expression patterns),提示miRNAs有可能作為參與調控基因表達的分子,因而具有重要意義。
第一個被確認的miRNA——線上蟲中首次發現的lin-4和let-7,可以通過部分互補結合到目的mRNA靶的3’非編碼區(3’UTRs),以一種未知方式誘發蛋白質翻譯抑制,進而抑制蛋白質合成,通過調控一組關鍵mRNAs的翻譯從而調控線蟲發育進程(reviewed in Pasquinelli 2002)。
bantam miRNA是第一個被發現有原癌基因作用的miRNA。除了lin-4、let-7,已知還有一些miRNAs可能參與在細胞分化和組織發育過程中起重要作用的基因的轉錄後調控,例如mir-14、mir-23 等。
在植物miRNAs的研究中有兩條線索提示miRNAs可能參與植物的發育過程。一是在carpel factory (car) 突變株中3個miRNAs的表達水平顯著下降。CARPEL FACTORY 是一個類似Dicer的酶,參與植物的發育,其缺失突變株表現為胚胎和葉片發育的缺陷。實驗結果提示這種缺陷是由於缺少miRNAs加工而造成的。多數的植物miRNAs在某些特定組織中高水平表達也提示他們可能參與了植物組織的發育。
對一部分miRNAs的研究分析提示:miRNAs參與生命過程中一系列的重要進程,包括早期發育(Reinhart 2000),細胞增殖,細胞凋亡,細胞死亡(Brennecke 2003),脂肪代謝(Xu 2003)和細胞分化(Kawasaki 2003)。此外,一個研究表明,2個miRNAs水平的下降和慢性淋巴細胞白血病之間的顯著相關,提示miRNAs和癌症之間可能有潛在的關係(Calin 2002)。
由於miRNAs存在的廣泛性和多樣性,提示miRNAs可能有非常廣泛多樣的生物功能。儘管對miRNA的研究還處於初級階段,據推測miRNAs在高級真核生物體內對基因表達的調控作用可能和轉錄因子一樣重要。有一種看法是:miRNAs可能代表在一個新發現的層次上的基因表達調控方式。
然而,大多數miRNAs的功能仍然是個謎。
MicroRNA的過表達
 micro RNA
micro RNAMicroRNA存在多種形式,最原始的是pri-miRNA ,長度大約為300-1000個鹼基pri-miRNA經過一次加工後,成為pre-miRNA 即microRNA前體,長度大約為70-90個鹼基;pre-miRNA再經過Dicer酶酶切後,成為長約20-24nt的成熟miRNA 。實際研究中,pre-miRNA套用最早,也最廣泛,目前很多商業化的MicroRNA庫都是pre-miRNA形式的。近幾年來,研究發現microRNA的雙臂對成熟miRNA的形成有著十分重要的作用,所以天然的pri-miRNA形式越來越多地被研究者採用。
MicroRNA的下調
化學合成的miRNA inhibitors ,用於下調目的細胞中的miRNA ,以實現loss-of function研究。
如果您需要進行長期、穩定的miRNA下調,則可以選用載體形式的miRNA inhibitor。其轉染效率高,下調效果好,可以實現對目的miRNA的長期、穩定的下調。
 micro RNA
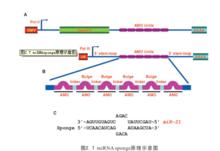
micro RNA載體形式的miRNA inhibitor,採用的方法如miRNA sponge法,這也是目前SCI文獻中用的較多的一種方法。
作用方式
microRNA-RISC對靶基因mRNA的作用主要取決於它與靶基因轉錄體序列互補的程度,有三種方式。
第一種是切斷靶基因的mRNA分子——miRNA與靶基因完全互補結合,作用方式和功能與siRNA非常相似,最後切割靶mRNA。在植物中,大部分miRNA都以這種方式,靶基因mRNA斷裂後,無poly(A)的分子的3‘ 端加上多個U並很快降解,含poly(A)的分子能穩定存在一段時間(如擬南芥miR-171)。在植物中目前有一個miRNA和3個潛在的目標靶基因完全互補(這些scarecrow 基因編碼潛在的轉錄因子),儘管還不清楚這些基因是否就是miRNA的目標靶,這仍是第一次發現miRNA 和其潛在的目標靶完全互補,也提示miRNA可能包含和siRNA類似的作用方式。
第二種是抑制靶基因的翻譯——作用時與靶基因不完全互補結合,進而阻遏翻譯而不影響mRNA的穩定性,這種miRNA是目前發現最多的種類(如線蟲lin-4)。而在植物中極少數的miRNA通過此方式來抑制靶基因。
第三種是結合抑制——具有以上兩種作用模式:當與靶基因互補結合時,直接靶向切割mRNA;當與靶基因不完全結合時,起調節基因表達的作用。
識別方法
多個研究小組採用生物化學結合是生物信息學的方法開展對miRNAs的研究工作。由於據推測都是由Dicer酶降解RNA得到的,21—23個鹼基大小、有5’端磷酸基和3’羥基的RNA片斷,有的實驗室採用改良的定向克隆方法來篩選具有相同特徵的小分子——篩選一定大小的RNA分子,連線到3’和5’的適配子(adapters),逆轉錄並通過PCR擴增、亞克隆並測序。miRNA前體在基因組上的定位和聚類是通過向基因組資料庫查詢進行。這個方法有助於判斷miRNAs是否是mRNAs、tRNAs、rRNAs等分子的降解產物。
有的實驗室通過一種RNA folding program ’mfold’ 來判斷C. elegans 和C. briggsae 之間的高度保守區域是否含有潛在的miRNA前體,然後用Northern Blots的方法來確定這些miRNAs是否真的表達了。
儘管有數百個miRNAs通過生化或者是生物信息學的方法被鑑別出來,已經鑑別出來的miRNAs只不過是滄海一粟,由於很多已經鑑別出來的miRNAs是從單個克隆中鑑別出來的,所以可以假設還有很多miRNAs在分離和鑑定過程中被“漏掉”了,測序工作還遠遠不夠。
siRNA
miRNA和siRNA之間的關係令人迷惑。從表面上說,一個是非編碼的單鏈小分子RNA,在進化上高度保守,通過翻譯抑制調控基因表達;另一個是針對編碼區的雙鏈小分子RNA,每個轉錄本都可能有很多個siRNAs,是通過降解目標靶,在轉錄後調控基因表達。由於每個mRNA模版可能產生很多個siRNAs,要給每個siRNA定一個基因的名字就很困難。miRNA是進化進程中高度保守的,因此給直向同源物一個同樣的名字可能有助於了解他們的功能,而給另一個物種中一段無關的序列一個同樣的名字就容易造成混亂。
然而,據推測miRNAs通常是由較大的(7090 nt)的莖環結構(髮夾結構)前體經Dicer酶切割得到的,而Dicer同樣負責將長雙鏈RNA切割為siRNA,而且二者的長度也差不多,同樣有調控基因表達功能。因而這兩類小分子RNA之間的關係格外令人關注。
兩個廣為人知的miRNA——線上蟲中首次發現的lin-4 和let-7,通過一種未知方式誘發蛋白質翻譯抑制從而抑制蛋白質合成。這種結合併不誘導mRNA靶的降解,就是說作為翻譯抑制子本身不影響對應mRNA的豐度,其原因據推測是由於miRNA和結合位點之間不完全互補。這就區別於siRNA的介導的mRNA的降解。但是其他一些miRNAs可能以類似siRNA的方式介導目的RNA的降解。實驗表明引入和let-7目的mRNA靶完全互補的miRNA會誘導mRNA靶的降解。還有實驗結果表明一些miRNA,包括在植物中發現的Scarecrow miRNA,能結合完全互補的mRNA鏈從而降解mRNA序列,抑制蛋白合成。這提示miRNAs可以和siRNAs一樣作用,這兩種小分子RNA作用通路可能有重疊的部分。這種重疊同樣提示siRNAs可能也有和miRNAs同樣的功能。
一個很有趣的實驗證實這個觀點:Doench和同事挑選一個已知在體內可以有效使CXCR4基因沉默的siRNA,然後在螢光素酶報告基因的3’端插入對應的CXCR4結合位點——其中一個拷貝是插入一個完全匹配的CXCR4結合位點,另一個拷貝插入4個只有3’和5’端匹配,而中間不同的CXCR4結合位點,這樣選定的siRNA就不能完全結合到這個結合位點——中間形成一個突起的不匹配的環。將這兩個拷貝轉入Hela細胞並用siRNA誘導基因沉默。結果很有趣——兩個實驗都錄得螢光素酶活性下降了超過10倍,RT-PCR和Northern分析證實,第一個實驗的螢光素酶轉錄本下降了超過10倍,這正是正常的siRNA介導的RNAi反應,目標靶mRNA降解導致表達水平的下降,而第二個實驗中螢光素酶轉錄本僅僅下降1.2倍,這種目的基因表達水平下看起來象源於miRNA介導的翻譯抑制降,而不是siRNA介導的影響mRNA的穩定性導致。實驗表明:siRNA可能以miRNA的方式作用於mRNA。實驗人員還進行了另一個實驗:改變第二個實驗中的不匹配環的鹼基序列看起來不影響抑制效果,但是siRNA和報告基因上的結合位點的匹配程度越高抑制效果越好,增加siRNA的量,抑制效果越好——這一點和siRNA抑制的情況一樣——唯一不同的是:完全匹配的結合位點(siRNA作用方式)可以單獨起作用而相互不影響,而增加不完全配對的結合位點(注意在第二個實驗中用了4個CXCR4結合位點)的個數對翻譯抑制有顯著的加乘作用。
在哺乳動物細胞中還沒有找到內源的siRNA,外源的siRNA介導的RNAi作用正是一種抵禦機制。而miRNAs則廣泛存在於哺乳動物細胞中,從理論上推測可能參與多種調控作用。這兩種小東西的作用機制和相互關係的本質就顯得更加撲朔迷離。如何在實驗中正確鑑定siRNA和miRNA,甚至是其他的小分子RNA都成為一個值得關注的問題。
待解決問題
miRNAs在多個物種中廣泛被發現,而且在進化上高度保守。這些“小玩意兒”留給我們一大堆謎團:miRNA的確切功能是什麼?它的目標靶是什麼?作用機制是什麼?也許需要對植物或者線蟲的基因組進行miRNAs突變株的篩選,在果蠅中可以用targeted-disruption缺失miRNA序列。對miRNA突變株伴隨的表型缺失進行研究,有助於解釋miRNAs的功能。
正如Phillip Zamore說的:“如果miRNAs在進化的進程中如此高度保守而沒有任何實際功能,那真是大自然拿科研人員開涮——而且是一個殘酷的玩笑”。
研究工具
隨著小分子RNA日益受到研究人員的重視,很多研究小分子RNA的新方法不斷推出。
分離
由於小分子RNA可能參與分化、發育、組織生長、脂肪代謝等生理過程,在不同的組織和發育階段的表達水平有所不同,進一步了解小分子RNA的生物功能需要確定其在各種生物樣品中的表達水平,因而需要一種精確的定量純化方法,從而得到可信的數據。
現行的RNA純化方法包括有機溶劑抽提+乙醇沉澱,或者是採用更加方便快捷的矽膠膜離心柱的方法來純化RNA。由於矽膠膜離心柱通常只富集較大分子的RNA(200nt以上),小分子RNA往往被淘汰掉,因而不適用於小分子RNA的分離純化。有機溶劑抽提能夠較好的保留小分子RNA,但是後繼的沉澱步驟比較費時費力。mirVana miRNA Isolation Kit是採用玻璃纖維濾膜離心柱(glass fiber filter,GFF),既能夠有效富集10mer以上的RNA分子,又能夠兼備離心柱快速離心純化的優點,是一個不錯的選擇。對於特別稀有的分子,由於需要分離大量RNA而導致高背景而降低靈敏度,還可以進一步富集10mer到200bp的小分子RNA來提高靈敏度。
探針製備
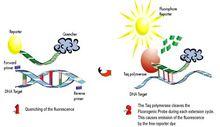
方法其實很簡單:只需要準備目的基因的一小段寡核苷酸序列,3’端另外增加8個和T7啟動子互補的鹼基,將這段寡核苷酸和T7啟動子引物退火,用Klenow大片斷補齊得到雙鏈的轉錄模版,然後用T7 RNA聚合酶、rNTP和標記物混合,體外轉錄得到標記的小分子RNA探針。這種方法可以快速製備各種標記(同位素、非放射性標記均可)的小於100nt的小分子RNA探針,適用於包括RPAs,Northerns 和原位雜交等各種方法檢測小分子核仁RNA( small nuclear RNA,snRNA),small interfering RNA (siRNA),,micro RNA (miRNA)和 mRNA。非放射性標記的探針還可以用於原位雜交研究miRNA或者mRNA在組織中的分布。
檢測
由於小分子RNA是一類很小的分子,部分小分子RNA表達水平可能很低,因而需要極為靈敏而定量的分析工具。由於其分子很小,用RT-PCR的方法來定量研究非常困難,多數研究人員採用Northern Blots——一種技術複雜而費力的方法來檢測小分子RNA的存在。傳統的Northern Blot的方法是是用探針檢測固相支持物(膜)上的目標分子,由於用探針檢測液相中的目標分子遠比檢測固相中的目標更為靈敏,生物通在這裡為大家推薦一種基於核酶保護分析方法改進的新方法——將同位素標記好的小分子RNA探針和待檢測樣品混合雜交,未雜交的RNA和多餘的探針用單鏈核酸酶消化,然後使核酸酶失活,並純化雜交的RNA分子,最後通過變性膠電泳放射自顯影檢測結果。這個基於液相雜交的新方法不但操作簡單而快速,而且靈敏度極高——可以半定量檢測少至10ng總RNA模版中的小分子RNA,或者說,可以檢測attomole (10-18 mol)級別的靶目標!靈敏度是Northern Blot的100倍。除此之外,研究人員還可以在同一個樣品中同時檢測多個小分子RNA和長的RNA模版。應該說,這個靈活巧妙的設計可以為從事小分子RNA實驗的研究人員帶來不少方便。
總而言之,無論是siRNA, miRNA, snRNA還是其他的小東西,小分子RNA研究的不斷深入將幫助我們揭示更多生命的奧秘。
功能分析
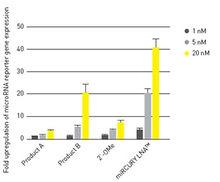 MicroRNA功能分析
MicroRNA功能分析miRNA 的上調可用於鑑定功能獲得表型;抑制或下調可以研究功能缺失表型。上調與下調的結合可用於鑑定被特定miRNA 調節的基因,以及特定miRNA 參與的細胞進程。
主套用包括:
◇miRNA 靶定位點的鑑定和驗證
◇篩選調節某個特定基因表達的miRNAs
◇篩選影響某個特定細胞進程的miRNAs
miRNA展望
miRNA在細胞分化,生物發育及疾病發生髮展過程中發揮巨大作用,越來越多的引起研究人員的關注。隨著對於miRNA作用機理的進一步的深入研究,以及利用最新的例如miRNA晶片等高通量的技術手段對於miRNA和疾病之間的關係進行研究,將會使人們對於高等真核生物基因表達調控的網路理解提高到一個新的水平。這也將使miRNA可能成為疾病診斷的新的生物學標記,還可能使得這一分子成為藥靶,或是模擬這一分子進行新藥研發,這將可能會給人類疾病的治療提供一種新的手段。
