
物種介紹
蕨類植物門——Pteridophyta
為維管束的孢子植物(也稱高等孢子植物),陸生、附生、少為水生,直立或少為纏繞攀援的多年生草本,或間為高大樹形;孢子體(即通常所謂綠色蕨類植物)照例有根、莖、葉的器官分化[松葉蕨(蘭)除外]。孢子體的形體在近代植物界中最為多種多樣。有大如喬木狀的,也有小僅達1厘米的,但絕大多數為中形多年生草本。孢子體生有多數孢子囊,內生孢子;最原始蕨類植物的孢子囊生於枝之頂端,有些生在特化的葉上或葉片上(囊托)成穗狀或圓錐狀囊序,有的生於孢子葉的邊緣,也有的聚生於枝頂成孢子葉(囊)球,而在絕大多數的種類則以各種形式生於孢子葉的下面,形成所謂孢子囊羣(堆)、或滿布葉之下面。孢子(有n染色體)有同孢和異孢之分:異孢型的,在孢子體(郎通常的植物體)上生大小兩種孢子葉:大孢子葉生大孢子囊,內生大孢子;小孢子葉生小孢子囊,內生小孢子。近代絕大多數的蕨類植物都屬於同孢型。孢子成熟後從孢子囊內以特種巧妙的機制(環帶)被散布出來,落地後萌發生長成為原葉體,叫做配子體。配子體的形體甚為簡單,為不分化的葉狀體、塊狀體或分叉的絲狀體等。在同一配子體上產生頸卵器和雄精器(雌雄同體),但在異孢型的蕨類植物,配子體更為簡化而有雌雄性之分(雌雄異體),雄配子體極小,不脫離小孢子壁,雌配子體遠較大,也不脫離大孢子壁。精子憑水為媒介,藉助於本身的纖毛運動,能和卵子行受精作用,產生配偶子,由此生長發育成為綠色孢子體(有2n染色體),也可叫作成長的蕨類植物,在它的葉體上產生孢子囊,內生孢子,孢子落地萌發又發育成原葉體。這樣,孢子世代的孢子體和配子世代的配子體相互交替一次,就完成蕨類植物的生活周期。
現代蕨類植物約有11,500多種,廣泛分布於世界各地,尤以熱帶和亞熱帶最為豐富。中國約有2 000種,它們大都喜生於溫暖陰濕的森林環境,成為森林植被中草本層的重要組成部分,不僅對森林的生長發育有著重大影響,同時可以作為敏感地反映環境條件的指示植物。
我國的蕨類植物是世界最豐富的一區,其中許多種類為藥用植物,還有一些作為蔬菜之用,另有一些是澱粉植物。最普遍利用的首推蕨菜的地下根狀莖。根據商業部初步統計,僅1958年,由於各地農民利用蕨根內的澱粉(俗稱蕨粉)製造各種食品的結果,為國家節約大量糧食。蕨類植物的枝葉青翠,姿態奇特,可供美化庭園,有引入入勝之感,而且許多種類為室內點綴的重要盆景。
特別可以指出,不少蕨類在工業生產上有很大意義,其中最重要的是石松科的許多種,它們的孢子(俗稱石松粉)為冶金工業上的優良脫模劑,可以提高鑄件的品質。由於蕨類植物的古老性,它們的化石和孢子為鑑定地層的一個重要指標。在自然科學領域內,蕨類植物,由於它在整個植物界的獨特位置——介於低等植物和高等植物之間——以及它的形態桔構的多樣性,常被當作研究生物進化和植物系統發育理論的重要對象之一。在這方面的科學研究工作,我國是一具備著優越條件的。
形態特徵
從橫切面看,莖的表皮層由外層加厚細胞組成,其內為皮層及中柱體。蕨類植物的中柱體較為複雜,隨科屬而不同,有時同一株植物隨不同發育年齡而有所改變,如桫欏的莖,其基部為原生中柱,中部為,頂部為網狀中柱。中柱的主要類型有:原生中柱,中柱中央分化為木質部,內層為韌皮部;管狀中柱,中柱中央為薄壁細胞組成的髓,向外為木質部和韌皮部,由於韌皮部的位置不同,又可分為外韌管狀中柱和雙韌管狀中柱,前者僅木質部外圍有一圈韌皮部,後者木質部的內外均有一圈韌皮部;網狀中柱,由管狀中柱分裂而成的一種,但維管束仍呈管狀排列;多環中柱,網狀中柱的各個維管束再次分裂成不規則的分散狀排列。和種子植物一樣,蕨類植物的莖的維管束由管胞、篩管、薄壁組織和內皮層組成,但無形成層,不能進行次生生長。在高大如喬木的種類中,其莖幹也只是由於薄壁細胞的增加而有限的加粗。
③葉:通常兼有營養和生殖兩種功能,即綠色的葉片在全部或部分羽片的下面一定部位,都可產生孢子囊,是謂葉片一型。有的科屬在同一植株上,先後長出兩種不同形狀的葉片,一種為正常的綠色營養葉或稱不育葉,另一種幼時為綠色,不久即長出孢子囊而失去綠色的孢子葉,又稱能育葉,這種類型叫葉片二型。少數種類,如紫萁屬(Osmunda)中,同一片葉上生有能育和不育兩種不同形狀的羽片,這叫羽片二型。
傳統上根據葉片不同把蕨類植物分為兩大類:小葉型蕨類,或稱擬蕨類,葉小,長寬僅數毫米(如石松、卷柏),或退化成鱗片狀(如松葉蕨),或為膜質(如木賊),僅水韭葉大而成線形,但和其他擬蕨一樣,無柄,都只有一條中肋而無完善的葉脈系統(據最新發現,熱帶的石松中有個別種具簡單的分枝葉脈),孢子囊生於葉腋;大葉型蕨類,即真蕨類,除槐葉的沉水葉變為鬚根狀,少數屬種的葉軸頂端延伸成鞭伏,能著地生根,以及部分科的植物葉軸能無限生長外,其餘都為正常葉。幼時通常拳卷,長大分葉柄和葉片兩部分,除單葉外,複葉的葉片有葉軸。按分裂的次數不同,而有一回、二回……羽狀複葉;羽片,一回小羽片、二回小羽片……;以及羽軸、一回小羽軸、二回小羽軸……之分(圖2),在一回羽狀複葉中,如果羽片邊緣分裂,則按分裂的深淺,稱之為葉片一回羽狀,羽片淺裂,或葉片一回羽狀,羽片深裂(達羽軸的狹翅或闊翅),習慣上也統稱之為二回羽狀淺裂,或二回羽狀深裂(達羽軸的狹翅或闊翅)。如果羽片裂達羽軸,裂片彼此分離,或形成小羽柄,則稱這種葉片為二回羽狀,其分離的裂片稱小羽片。依此類推,直至末回裂片。
蕨類植物的葉片結構除少數屬種有柵狀組織和海綿組織分化外,一般僅有海綿狀、多少有空隙的葉肉層,有的甚至連葉肉層也沒有,光合作用全靠表皮來完成。通常上表皮不具氣孔或有極少氣孔,下表皮的氣孔類型很多,因屬種而不同。但也有同一種葉片下表面具好幾種氣孔型的。 蕨類植物的葉脈比較簡單,絕大多數是分離的,少有簡單網結,網眼無內藏小脈或僅有少數內藏小脈。
有些屬種,葉片側脈的分出和小羽片的分出一樣有上先出和下先出之分。先從各回羽軸上側分出的小羽片,叫小羽片上先出,反之叫下先出。同樣,先從主脈上側分出的側脈叫上先出脈,又叫上行脈序,反之叫下行脈序。這一特徵,在某些屬種中穩定而有鑑別價值。
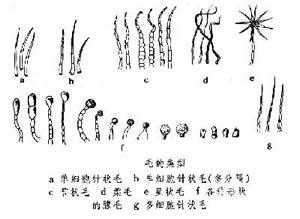 毛的類型
毛的類型 ④毛被:蕨類植物的毛被相當複雜,分毛和鱗片兩大類,或調節葉面空氣濕度,或保護孢子囊群。有些腺毛 可能和代謝有關,分泌蠟質粉末的腺毛,有保持水分的功能。
⑤孢子囊:無性繁殖器官,產生在葉片的一定部位,由於它的構造和形成的不同,可分為厚囊和薄囊兩大類,前者的囊殼由多層細胞組成,後者的囊殼僅 1層。在發育上,兩者的原始細胞都經橫斷分裂成上下兩枚細胞,所不同的是厚囊蕨類的上枚為囊殼細胞,下枚為孢原細胞,而薄囊蕨類的下枚細胞為柄細胞,以後發育成孢子囊柄,上枚細胞經魚鱗式斜分裂成四面體,再分裂為囊殼和孢原細胞。因此厚囊蕨的孢子囊壁厚而無柄,薄囊蕨的孢子囊壁薄而有柄。
⑥環帶:薄囊蕨類孢子囊釋放孢子的機構。由數個到數十個U形加厚細胞和多個扁平的薄壁細胞(包括唇細胞)組成,環繞囊殼。當孢子成熟時,由於環帶的U形細胞失水收縮而產生的拉力,孢子囊的唇細胞被拉開,而將孢子彈出。環帶的有無和存在部位,常和進化有一定關係。通常較原始的無環帶,如厚囊蕨,或僅有不發達的環帶(如紫萁),逐漸發展為頂生環帶(如海金沙)、橫生中部(如芒萁)、斜行環帶(如瘤足蕨)和進化類型的縱型而中斷於囊柄的水龍骨型環帶。
⑦孢子囊群:擬蕨類的孢子囊常近軸而單生於葉腋,其中有些屬種,如石松、卷柏的一些種和木賊屬。長孢子葉的枝縮短而形成或長或短的孢子囊穗。真蕨類孢子囊通常遠軸而生於葉片下面(原始類群的生於葉緣)。它們通常多數聚生在位於葉脈的表皮分化形成的囊托上,而呈各種形狀的孢子囊群(sorus),有些厚囊蕨類的孢子葉常形成穗狀或圓錐花序狀。較原始類群的孢子囊群的孢子囊數目少而同時發育,發展為從內向外逐漸發育,而進化類型的孢子囊群中的孢子囊為混合發育,即一個囊群里有的發育早,有的發育晚或敗育,這在有些種里,敗育的孢子囊就成為一種隔絲。
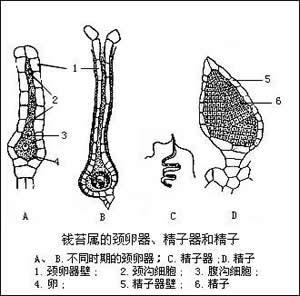 頸卵器
頸卵器 ⑧隔絲:又稱夾絲。是一種毛狀不育器官,混在孢子囊群中起內外保護作用。一般都有長柄,頂部呈各種形 狀,覆蓋幼孢子囊群,如石韋的星狀毛隔絲,瓦韋的盾狀隔絲等。也有許多隔絲。雖有長柄,但不伸出囊群之外,夾在囊群中起內保護作用。 ⑨囊群蓋:在薄囊蕨綱中異型孢類的孢子囊有一個由葉片特化形成的外殼,包被在大、小孢子囊外面,稱之為大、小孢子果,而同型孢類的孢子囊群大都有囊群蓋。它主要由葉片表皮細胞分化而來,初時綠色,老時黃褐色,用以保護孢子囊群。囊群蓋由於在囊托上形成的位置不同而有上位和下位之分。囊群蓋環繞囊托的基部生出,把孢子囊包在蓋內的叫囊群蓋下位,或叫上位囊群,而囊群蓋生於托的頂端或一側,從上面覆蓋孢子囊群的叫囊群蓋上位,有的雖生於一側,但被成熟的孢子囊壓在下面,有如半下位。還有些孢子囊群靠近葉邊生的,其葉邊往往多少特化變質而向下反卷覆蓋孢子囊群,這種稱為假蓋。而有些種類的蓋則以毛或鱗片形式出現(如岩蕨屬中),囊群蓋的形式多樣,為鑑別屬的重要特徵。
區別
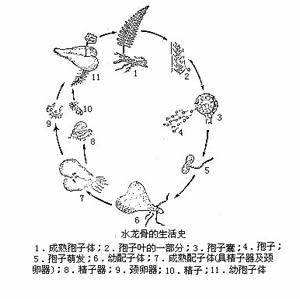
通常肉眼所見到的綠色蕨類植物即是它的孢子體,孢子體上產生孢子囊每一個孢子囊內產生大量的、肉眼看不清的、直徑10-100微米的孢子。孢子比重小,懸浮於空氣中,隨氣流或水流漂移到各處,在適當的環境中萌發,長出微水的綠色(少數不具葉綠素,不能自養)葉狀體──配子體(原葉體)。除部分擬蕨類和水生真蕨類孢子體產生大、小兩種孢子,萌發後成長的配子體有雌雄之分外,絕大多數蕨類都屬於同型孢類,它們的孢子無大小之分,並在孢子萌發後產生的配子體上的不同部位,產生頸卵器和藏精器。並由此分別產生雌配子——卵子和雄配子——精子,雌雄配子結合後形成合子,然後發育成胚,再成長為常見的綠色孢子體。這個過程就是蕨類植物特有的世代交替生活史。在整個生活史中,常因環境等條件的影響而改變其中某一過程,導致一些不正規的繁殖。常見的為無配子生殖和無孢子生殖。
一般特徵
特徵
 蕨類植物
蕨類植物 植物體已有真正的根、莖、葉和維管組織的分化。已屬維管植物的範疇。木質部只有管胞、韌皮部只有篩管或篩胞,沒有伴胞,不開花、不產生種子,主要靠孢子進行繁殖,仍屬孢子植物。生活史中有明顯的世代交替現象,孢子體世代占優勢。
配子體弱小,生活期較短,稱原葉體。孢子體和配子體均為獨立生活的植物體。習見植物體為孢子體,一般為多年生草本,少數種類為高大的喬木,如生活在熱帶的樹蕨,高可達20米。根通常為須狀不定根。莖多為地下橫臥的根狀莖,少數種類具有地上直立或匍匐的氣生莖。葉有單葉和複葉之分,葉形變化很大。有些蕨類植物,同一植物體上的葉可區分為形態和功能各異的孢子葉和營養葉(即異形葉)。孢子葉背面,邊緣或葉腋內可產生孢子囊,在孢子囊內形成孢子,以此進行繁殖,故又稱能育葉。營養葉僅有光合作用功能,不產生孢子囊和孢子,故又稱不育葉。
光合作用
 蕨
蕨 一般蕨類植物的葉子兼具進行光合作用製造有機養料和產生孢子進行 繁殖的功能(即同型葉)。蕨類植物是最古老的陸生植物。在生物發展史上,已有35000萬年到27000萬年的泥盆紀晚期到石炭紀時期,是蕨類最繁盛的時期,為當時地球上的主要植物類群,高大的鱗木、封印木、蘆木和樹蕨等共同組成了古代的沼澤森林。二疊紀末開始,蕨類植物大量絕滅,其遺體埋藏地下,形成煤層。
生長環境
大多為土生、石生或附生,少數為濕生或水生。喜陰濕溫暖的環境。高山、平原、森林、草地、溪溝、岩隙和沼澤中,都有蕨類植物生活。
分布範圍
 蕨類植物——桫欏
蕨類植物——桫欏 現存蕨類植物約12000種,廣泛分布在世界各地。尤以熱帶、亞熱帶地區種類繁多。我國約有2400種,主要分布在長江以南各省區。對蕨類植物的系統分類,分類學家的觀點尚不一致。我國植物學家秦仁昌1978年提出的觀點,被認為是更接近真正系統發育的較新的分類系體。據此,將蕨類植物分成5個亞門,即:松葉蕨亞門(Psilophytina)、楔葉蕨亞門(Sphenophytina)、石松亞門(Lycophytina)、水韭亞門(Isoephytina)和真蕨亞門(Filicophytina)。
生長過程
配子體
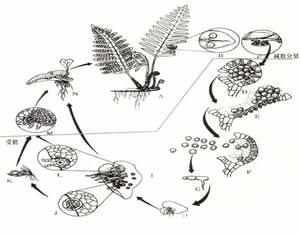 蕨類植物生長史(真蕨綱為例)
蕨類植物生長史(真蕨綱為例) 配 子體 有性世代的簡單植物體,由孢子萌發而來。孢子萌發後形成幾個細胞的簡單絲狀體,經過頂細胞和側生分生組織的細胞分裂形成片狀體,或稱細胞片,再發育為成熟的通常背腹扁平的心形原葉體,原葉體靠近尖端下面生有假根,藉以固定和吸收水分,凹缺處為生長點,雌器(頸卵器archegonium)生長在附近,其外形大而似瓶狀,腹部埋在組織內,頸部彎向雄器所在部位,口部突出。雄器(精子器antheridium)呈突起狀或圓球形,有柄或無柄,生長在假根附近,通常它的壁由 3個細胞:莖細胞、環細胞和帽細胞(如果帽細胞分裂,則為四個)組成,進化類型的由更多的細胞組成。每個頸卵器含一雌配子——卵子,每個精子器含多個雄配子──精子,精子頂部有纖毛。配子體這樣的結構,巧妙地利用了貼近地面,易於獲得水分的特點。通常雌配子器的發育晚於雄配子,成熟後除卵細胞外,頸卵器內其他細胞被分泌的有機酸溶解變成膠質而溢出。當原葉體下面有一層薄水層時,精子器的帽脫開(進化類型的為孔裂),精子被釋放,通過纖毛運動和受頸卵器口溢出膠質的化學感應而被吸引到頸卵器中,和卵子結合而產生合子,合子萌發後形成胚,再發育成常見的孢子體。 配子體通常壽命很短,在孢子長出後不久即死去,這是由於原葉體結構簡單,無維管組織。但也有生活期長的,如翠蕨Anogramma,在孢子體長出幾片葉後,原葉體仍然可見。有些種的配子體可生活4-5年。這些生活期長的配子體,通常較大而厚,出現在較原始的類群。
孢子體
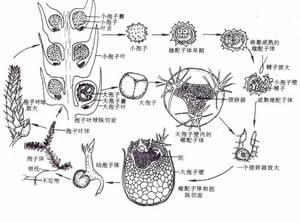 卷柏屬生長史
卷柏屬生長史 孢子體 無性世代植物體,具根、莖、葉和輸導系統的分化。
①根:蕨類植物只有不定根而無主根,通常叢生於直立的根狀莖末端,或散生在橫走根狀莖腹面,藉以固定根狀莖。根內具原生中柱,無次生生長,不能加粗,表面生有根毛,吸收水分和營養物質。
②莖:除桫欏科和蚌殼蕨科的部分種類具高大喬木狀直立莖外,絕大多數蕨類植物的莖都貼近地面稱根狀莖,或短而直立,斜升,或細長橫走,少數為纏繞藤本,極少數呈灌木狀。其頂端通常被毛或鱗片,有的種類還被葉柄殘莖組成的防護罩所保護。
分類系統
簡介
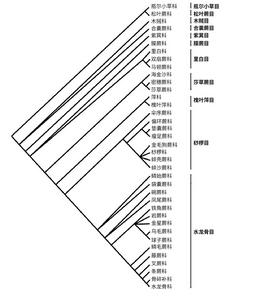 蕨類植物分類
蕨類植物分類 1940年中國蕨類植物學家秦仁昌將經典的十分混雜的水龍骨科(實際上是薄囊蕨綱的絕大部分)劃分為33科,促進了對蕨類植物系統的廣泛探討。世界上現有蕨類分類系統不下10個,大體上有兩類:其一是以美國E.B.科普蘭(1947)為代表的系統,其次是秦仁昌 (1940,1954,1957,1978)為代表的系統。各國學者根據各自所掌握的材料和觀點而有所增減或變化,總的說來,在絕大多數屬的劃分上,國際蕨類植物學界分歧性不大,而在科,特別是科以上等級排列還莫衷一是。1940年秦仁昌在將水龍骨科分為33科的同時,提出了5條進化乾,即雙扇蕨類、桫欏類、蚌殼蕨類和兩個絕滅祖先類,表示出各幹上科的關係。其後許多學者提出系統樹的構想,但由於這些系統樹各自根據單方面材料,如印度學者B.K.奈亞爾等主要根據形態解剖方面材料,美國W.H.小華格納等主要根據孢子囊方面材料,義大利的 R.E.G.皮基·塞爾莫利則偏重於染色體的材料等而過於簡單化,有的雖有綜合考慮,但例證不多,特別是在化石證據不足,生物系統學研究也還沒有普遍開展的情況下要提出一個比較自然的系統樹,還有待於全世界的共同努力。
分類
(按秦仁昌系統排列各科)
蕨類植物門 Pteridophyta
松葉蕨亞門 Psilophytina
也叫裸蕨類,是原始的陸生植物類群。孢子體僅有假根,氣生莖二叉分枝,葉為小型葉,無葉脈或僅有單一葉脈,孢子囊2~3枚聚生於枝端或葉腋,孢子同型。配子體雌雄同株,生地下,無葉綠體。大多已絕跡,現存僅松葉蕨目Psilotales,包括2科:松葉蕨科Psilotaceae和梅溪蕨科Tmesipteridaceae,共3屬3種。其中梅溪蕨科僅1種,僅見於大洋洲;松葉蕨科有2種,我國僅松葉蕨Psilotum nudum,產於西南、華南及華東地區。
松葉蕨目 Psilotales
01.松葉蕨科 Psilotaceae
石松亞門 Lycophytina
莖幹大多為二叉分歧,葉極小,有葉舌或無,螺旋狀排列或4列互動對生,但都只有單一不分枝的維管束。孢子囊生於孢子葉腋,有同型(石松)和異型(卷柏)之分。現存僅石松目Lycopodiales和卷柏目Selagineales,前者包括3科:石杉科Huperziaceae、石松科Lycopodiaceae、石蔥科Phylloglossaceae,後者僅卷柏科Selaginellaceae,共約1000種。常見如卷柏Selaginella tamariscina、蛇足石杉Huperzia serrata、石松Lycopodiastrum japonicum、燈籠草Palhinhaea cernua、翠雲草Selaginella uncinata等。多產於熱帶,有些廣泛分布於溫帶,通常生於林下或附生樹幹,部分生於乾旱的石壁。其中石松孢子在工業上稱石松粉,為鑄造工業的優良脫膜劑,或入藥。
石松目 Lycopodiales
02.石杉科 Huperziaceae
03.石松科 Lycopodiaceae
卷柏目 Selaginellales
04.卷柏科 Selaginellaceae
水韭亞門 Isoephytina
水生草本,有根、莖葉的分化;具根托(屬於莖),先端生不定根;莖塊莖狀,原生中柱,有次生生長結構;葉條形叢生,具葉舌;孢子葉有大小之分,孢子異型;配子體極度簡化,有雌雄配子體之分。僅1目1科1屬約60餘種,即水韭目Isoetales水韭科Isoetaceae水韭屬Isoetes。我國僅3種:中華水韭Isoetes sinensis僅產於我國長江下游地區,可能已滅絕;寬葉水韭Isoetes japonica產於雲南、日本及朝鮮;台灣水韭Isoetes taiwanensis特產於台灣。生於沼澤、溝塘淤泥中,由於人為的環境破壞,已列為國家級保護植物。
水韭目 Isoetales
05.水韭科 Isoetaceae
楔葉亞門 Sphenophytina
植物體有根、莖葉的分化;有不定根;莖有節和節間之分,有時生塊狀莖,管狀中柱到具節中柱,木質部內始式;葉小型,不發達,輪生成鞘狀;孢子葉又稱孢囊柄,孢子同型或異型;配子體具背腹性。出現於泥盆紀,繁盛於石炭紀。現存僅1目1科1屬,即木賊目Equisetales木賊科Equisetaceae木賊屬Equisetu,共29種。常見如問荊Equisetum arvense和木賊Equisetum hiemale,前者有營養莖和生殖莖(孢子莖)之分,後者則無,另如節節草Equisetum ramosissimum、筆管草Hippochaete debile。多生於田邊、溪溝邊、路邊和林緣等陰濕地方。
木賊目 Equisetales
06.木賊科 Equisetaceae
真蕨亞門 Filicophytina
為現今最繁茂的蕨類植物,約有1萬種以上,廣布全世界;我國近2000種,廣布全國。孢子體發達。莖除了樹蕨外,均為根狀莖,有各式中柱。木質部主要成分為管胞,少數具導管。葉為大型葉,幼葉拳卷狀,長大後平展,分化為葉柄和葉片二部分。葉片為單葉或一至多回羽狀分裂或複葉。孢子囊常聚集成孢子囊群生於葉邊緣或背面,也有的生於特化了的孢子葉上,有或無囊群蓋,孢子同型,一些水生真蕨的孢子囊則生於特化的孢子果內,孢子異型。 配子體為心臟形的葉狀體,長寬一般不超過1厘米,綠色,有假根。精子器和頸卵器均生於腹面。根據孢子囊的發育方式、結構及著生位置等,可分為原始薄囊蕨綱Protolyptosporangiopsida、厚囊蕨綱Eusporangiopsida和薄囊蕨綱Leptosporangiopsida。如蕨Pteridium aquilinum、海金沙Lygodium japonicum等。
厚囊蕨綱 Eusporangiopsida
瓶爾小草目 Ophioglossales
07.七指蕨科 Helminthostachyaceae
08.陰地蕨科 Botrychiaceae
09.瓶爾小草科(箭蕨科) Ophioglossaceae
蓮座蕨目 Marattiales
10.合囊蕨科 Marattiaceae
11.蓮座蕨科 Angiopteridaceae
12.天星蕨科 Christenseniaceae
原始薄囊蕨綱 Protoleptosporangiopsida
紫萁目 Osmundales
13.紫萁科 Osmundaceae
薄囊蕨綱 Leptosporangiopsida
水龍骨目(真蕨目) Polypodiales(Filicales)
14.瘤足蕨科 Plagiogyriaceae
15.裏白科 Gleicheniaceae
16.莎草蕨科 Schizaeaceae
17.海金沙科 Lygodiaceae
18.膜蕨科 Hymenophyllaceae
19.蚌殼蕨科 Dicksoniaceae
20.桫欏科 Cyatheaceae
21.稀子蕨科 Monachosoraceae
22.碗蕨科 Dennstaedtiaceae
23.鱗始蕨科 Lindsaeaceae
24.竹葉蕨科 Taenitidaceae
25.姬蕨科 Hypolepidaceae
26.蕨科 Pteridiaceae
27.鳳尾蕨科 Pteridaceae
28.鹵蕨科 Acrostichaceae
29.光葉藤蕨科 Stenochlaenaceae
30.中國蕨科 Sinopteridaceae
31.鐵線蕨科 Adiantaceae
32.裸子蕨科 Hemionitidaceae
33.水蕨科 Parkeriaceae(Ceratopteridaceae)
34.車前蕨科 Antrophyaceae
35.書帶蕨科 Vittariaceae
36.蹄蓋蕨科 Athyriaceae
37.腫足蕨科 Hypodematiaceae
38.金星蕨科 Thelypteridaceae
39.鐵角蕨科 Aspleniaceae
40.睫毛蕨科 Pleurosoriopsidaceae
41.球子蕨科 Onocleaceae
42.岩蕨科 Woodsiaceae
43.烏毛蕨科 Blechnaceae
44.球蓋蕨科 Peranemaceae
45.鱗毛蕨科 Dryopteridaceae
46.叉蕨科 Aspidiaceae
47.實蕨科 Bolbitidaceae
48.藤蕨科 Lomariopsidaceae
49.舌蕨科 Elaphoglossaceae
50.腎蕨科 Nephrolepidaceae
51.條蕨科 Oleandraceae
52.骨碎補科 Davalliaceae
53.雨蕨科 Gymnogrammitidaceae
54.雙扇蕨科 Dipteridaceae
55.燕尾蕨科 Cheiropleuriaceae
56.水龍骨科 Polypodiaceae
57.槲蕨科 Drynariaceae
58.鹿角蕨科 Platyceriaceae
59.禾葉蕨科 Grammitidaceae
60.劍蕨科 Loxogrammaceae
苹目 Marsileales
61.苹科 Marsileaceae
槐葉苹目 Salviniales
62.槐葉苹科 Salviniaceae
63.滿江紅科 Azollaceae
下級分類
| 門 | 亞門 | 綱 | 目 |
| 蕨類植物門 | 松葉蕨亞門 | 松葉蕨綱 | 松葉蕨目 |
| 石松亞門 | 石松綱 | 石松目 | |
| 卷柏目 | |||
| 水韭亞門 | 水韭綱 | 水韭目 | |
| 楔葉亞門 | 木賊綱 | 木賊目 | |
| 真蕨亞門 | 厚囊蕨綱 | 瓶爾小草目 | |
| 蓮座蕨目 | |||
| 原始薄囊蕨綱 | 紫萁目 | ||
| 薄囊蕨綱 | 水龍骨目(真蕨目) | ||
| 苹目 | |||
| 槐葉苹目 |
繁殖類型
無配子生殖
 無配子生殖
無配子生殖 無 配子生殖 生活史中,配子體不產生配子而直接長出新孢子體。但這種配子體或是由僅經過有絲分裂產生的2n孢子萌發而來,或是孢子體先產生復二倍體,再經過減數分裂產生2n的孢子萌發而來。這種現象在熱帶產的同型孢類中和石松的一些種中很常見。
無孢子生殖
在孢子體上直接產生配子體,在自然界這是多倍體形成的一條途徑。在實驗條件下,控制培養基的成分和比例常可誘導出這兩種現象。有人認為這和能量有關,能量充裕出現前者,而能量貧乏則出現後者。
此外,在自然界營養繁殖的現象也很普遍。在孢子體的一些部位(如葉軸、羽軸或葉脈)能產生芽孢。還有的葉軸頂端常延伸成鞭狀,能著地生根,產生新株。
蕨類植物的孢子體和種子植物一樣具根、莖、葉和輸導系統的分化,能適應陸地生活,獲得更多的營養,從而產生和種子一樣起生殖和傳播作用的孢子。雖然孢子沒有種子在結構機能上的許多進步特徵,但由於它的量大,體積小,能被風吹到很遠地方,總有一些會遇到適宜的環境而萌發,因此,在某種意義上說更便於繁殖。
作用因素
形態結構起決定性作用
種子植物生活史的有性過程是在子房中度過的,而蕨類植物則和苔蘚植物一樣,配子體能獨立生活,也就是說有明顯的世代交替。所不同的是苔蘚植物是配子體世代占優勢,孢子體寄生在配子體上;而蕨類植物是孢子體占優勢,分別獨立生活。這一生活史特點表明蕨類植物是介於苔蘚和種子植物之間的一群植物。它的形態結構決定了它既是高等孢子植物或頸卵器植物,又是原始的維管束植物。
主要價值
豐富能源
 蕨類植物
蕨類植物 除古蕨類遺體在地層中形成的煤為人類提供了豐富的能源外,還有多方面經濟價值。多種蕨類可作藥用。如海金沙可治尿道感染、尿道結石;骨碎補能堅骨補腎、活血止痛;用卷柏外敷治刀傷出血;用貫眾治蟲積腹痛和流感;鱗毛蕨及其近緣種的根狀莖煎湯,為治療牛羊的肝蛭病的特效藥。蕨、菜蕨、水蕨、紫萁及觀音蓮座等都可食用,許多種蕨的根狀莖中富含澱粉,稱蕨粉或山粉,不但可食,還可作釀酒的原料。石松的孢子稱石松子粉,含有大量油脂,可作冶金工業上的優良脫模劑,使鑄件表面光滑,減少砂眼。木賊的莖含矽質較多,可作木器和金屬的磨光劑。滿江紅屬蕨類通過與固氮藍藻共生,能從空氣中吸取和積累大量的氮,既是優質的綠肥,又是豬、鴨等畜禽的良好飼料。
觀賞價值
蕨類植物的生活對外界環境條件的反應具有高度的敏感性,不少種類可作為指示植物。如卷柏、石韋、鐵線蕨是鈣質土的指示植物,狗脊、芒萁、石松等是酸性土的指示植物,桫欏與地耳蕨屬的生長,指示熱帶和亞熱帶的氣候。蕨類植物枝葉青翠,形態奇特優雅,常在庭院、溫室栽培或製作成盆景,具有較高的觀賞價值。
 蕨類植物
蕨類植物 