 遺傳作圖
遺傳作圖形態學標記
 雄果蠅的染色體組圖解
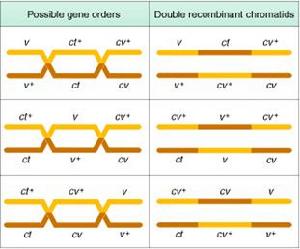
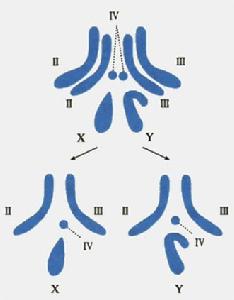
雄果蠅的染色體組圖解最初的遺傳學圖是在20世紀初對果蠅等生物構建的,使用基因作為際記:許多年之後人們才認識到基因是DNA分子的片段。而在當時,基因被認為是能將可遺傳的性狀從親子傳遞到後代的抽象實體一個遺傳性狀必須以兩種替換形式或表型(phenotype)存在才能用於遺傳學分析。如孟德爾首先研究的豌豆莖的高或矮。每種表型是由相應基因的不同等位基因(allele)所決定的。起切只有那些能通過視覺區分的基因表型能用於研究。比如,第一張果蠅遺傳圖顯示了負責身體顏色、眼睛顏色、翅膀形態等基因的位置,這些表型都可在低倍顯微鏡下或肉眼觀察果蠅而看到。
早期尚覺得這種方法很精細,但遺傳學家們很快就發現,只有有限的幾種可見表型的遺傳可用於研究,而在許多情況下,由於不止一個基因影響一個物理特徵,分析起來並不大容易。例如,到1922年,有超過50個基因被定位在4條果蠅染色體上、而其中9個基因負責眼睛的顏色,想在此領域有所貢獻的每一個初涉者必須首先學會辨別果蠅眼睛的顏色是紅、淡紅、朱紅、朽榴石色、康乃馨色、肉桂色、深褐色、猩紅或深紅色。因此,為了使基因圖更加全而,有必要找到一些比可見的性狀更多、更明確而且更簡單的性狀。
生化標記
 血清蛋白
血清蛋白以上問題的解決方案之一是套用生物化學方法來區分表型。這對於微生物與人類這兩種生物尤為重要,細菌與酵母等微生物只有為數很少的可見性狀,因此這類生物的基因作圖只能依賴於少數的生化表型。人類雖然有可見的性狀持征,但以血液分型為代表的生化表型研究從20世紀20午代就開始了。
血液分型研究不僅包括如ABO系統的標準血型,還有血清蛋白以及人類白細胞抗原〔HLA系統)等免疫蛋白的等位基因可變體。這些標記相對於可見表型的一個巨人優點是其相關基因往往為復等位基因(multiple allele)。例如,HLA—DRBI基因至少有59個等位基因,而HLA- B至少有60個。這正是與人類作圖相關的。與在果蠅或小鼠等生物中建立的雜交實驗不同,人類基因遺傳的數據只能通過檢查—個家族中各成員的表型來獲得。如果對所研究的基因而言,所有的家族成員都為純合子,就得不到有用的信息。這對於只有兩個等位基因的基因較常見,因為婚配可偶然地發生於同一個等位基因純合子個體之間。而當所研究的基因有60個而不是2個等位基因時,這就很少見了。
簡單的說:形態標記和生化標記的局限性在於多態性太低,可標記的位點太少。這是他們共同的致命弱點。
DNA分子標記
DNA 分子標記大多是以DNA片段電泳譜帶形式表現的。依其遺傳特性可分為顯性和共顯性標記2種;依多態性檢測手段可分為以Southern雜交技術為核心的分子標記和以PCR技術為核心的分子標記;根據在基因組中出現的頻率,又可分為低拷貝序列和重複序列標記。
(1)限制性片段多態性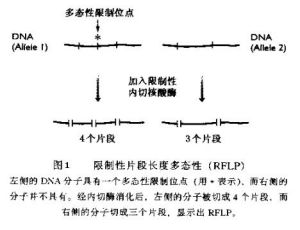 限制性片段長度多態性
限制性片段長度多態性(2)隨機擴增片段長度多態性標記(Random Amplified Polymorphic DNA ,RAPD)
RAPD技術是由Williams等首先創立的一種DNA分子標記技術,利用單一的10個鹼基寡核苷酸作為引物,對基因組DNA進行PCR擴增。經瓊脂糖凝膠電泳來檢測DNA序列多態性。
(3)擴增片段長度多態性(Amplified Fragment Length Polymorphism ,AFLP)
AFLP是Zeabeau 等(1993)發明的一項技術,它既有RFLP的可靠性,又有RAPD的方便性。其基本原理是通過PCR 擴增基因組DNA片段,擴增產物的變性聚丙烯醯胺電泳顯示擴增片段長度多態性,其中引物=接頭+酶切位點+2~3個核苷酸。
AFLP技術分析流程:(1)DNA 模板製備;(2)提取樣本DNA 經濃度和質量檢測後,一般採用雙酶(EcoRI和MSEI或PstI或TaqI)酶切, 在基因組DNA上產生低頻和高頻切口;(3)選擇性擴增酶切片段。酶切後,限制性片段在T4連線酶作用下與特定接頭連線,形成帶有接頭的特異性片段;(4)PCR前擴增,一般用帶一個選擇性引物進行預擴增,反應條件與常規PCR反應基本一致;(5)利用放射性同位素標記或螢光標記PCR 引物;(6)在Taq聚合酶作用下完成94 ℃變性30s ,65 ℃淬火30s,72 ℃延伸60s,PCR擴增36個循環;(7)PCR產物在含尿素聚丙烯醯胺上電泳;(8)將電泳後的凝膠轉移到吸附濾紙上,經乾膠儀進行乾膠處理;(9)在X光片上感光,數日後沖洗膠片並進行結果分析。
AFLP反應起始一般高溫復性(一般65 ℃),因此只有那些與3’端嚴格配對的片段才能得到擴增,選擇性很強。實驗結果穩定,重複性好,呈典型的孟德爾式遺傳,每個AFLP可以獲得50~100條譜帶信息,多態性強。
(4)微衛星(Microsatellite)
微衛星是指以幾個(1~6 bp) 核苷酸為單位,多次串聯重複序列,也稱之為簡單重複序列(single sequence repeats ,SSR) 、短串聯重複序列(short tandem repeats ,STR)或簡單序列長度多態性(single sequence length polymorphism ,SSL P) 。廣泛分布於真核生物基因組中,大約每隔10~50bp就有一個微衛星,由於重複次數和重複程度的不完全而造成每一個位點的多態性。
(5)單核苷酸多態性(Single Nucleotide Polymorphism,SNP)
基因組中存在單個的點突變,且數量極大,有些也對產生RFLP,但許多並個能。這是囪為它們所處的序列不能被限制件內切核酸酶所識別。在人類基因組中,據認為有200000個以上的SNP(single nucleotidc polymorphism)位乾基因內.而且有更多的SNP位於非基因的DNA中。
每個SNP只有兩個等位基因,所以這些標記在人類繪製遺傳圖譜方面又與RFLP同樣的缺點:對於一個SNP,很可
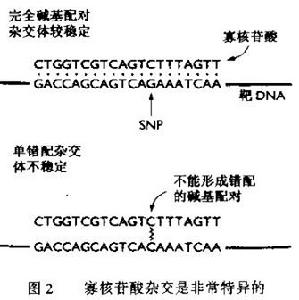 寡核苷酸雜交是非常特異的
寡核苷酸雜交是非常特異的DNA晶片(DNA chip)技術
DNA晶片是一塊面積為2cm平方或更小的矽片,以高密度排列方式攜帶有許多不同的寡核苷酸。待測DNA用螢光標記後加到晶片的表面。用螢光顯微鏡觀察雜交情況,顯示螢光信號的位置即表示該處的寡核苷酸與待測DNA發生了雜交。因此一個實驗中可以量化許多SNP。
動態等位基因特異的雜交(dynamic allele-specific hybridization, DASH)在這種技術中,雜交在溶液中進行,比如在96孔微量滴定板的一個孔中進行G螢光標記只能與雙鏈DNA結合,因此只有發生了雜交才能檢測到信號。開始,雜交在允許有錯配的條件下進行。在這個階段,無論待測DNA含有哪一種SNP等位基因,寡核苷酸都能與之雜交。由於錯配的雜交產物不如完全雜交產物穩定,故在較低溫度解鏈.因此可以通過升高溫度來區分等位基因。這樣就可以從使雜交依賴性螢光信號消失的溫度來判定待測DNA中存在哪一種等位基因。
