
基本內容
名稱
中文名稱( Chinese )→虛骨龍
其它中文名稱( Chinese[OTHER] )→ ...
 虛骨龍
虛骨龍拉丁文學名( Name )→ coelurus
發音( Pronounced )→ see-LURE-us
含義( Meaning )→ 尾巴中空的龍
屬種
目( Order )→ Saurischia 蜥臀目
亞目( Suborder)→ Theropoda獸腳亞目
類( Infraorder)→ Coelosauria 虛骨龍類[未定]
科( Family)→ Coeluridae虛骨龍科[未定]
屬( Genus )→ Chuandongocoelurus 虛骨龍
 虛骨龍
虛骨龍模式種( TYPE SPECIES )→ C. primitirus 原始虛骨龍
其它種( OTHER SPECIES )→ ...
錯誤歸屬種( MISASSIGNED SPECIES )→ ...
時代( Period )→ 侏羅紀晚期
分布( Found in )→ 亞洲及北美洲
發現地層( Stratum )→ ...
食性( Diet )→ 肉食
典型狀態( Length up to )→ 長2-3米
推測體重( Mass )→ ...
按照現代新的分類觀點,虛骨龍類也已列入聯尾龍類,其中包括鳥的祖先。它在進化上的顯著特徵是:在齶部的外孔,後肢比前肢約長一半。它主要分布於亞洲及北美洲。有許多不同的分類法將虛骨龍分成各種類群,這裡我們只介紹最重要的似鳥龍類和跑龍類。
歷史資料
 虛骨龍
虛骨龍奧塞內爾·查利斯·馬什在1884年繪製的腔骨龍脊椎圖。虛骨龍是在1879年由美國古生物學家與自然學家奧塞內爾·查利斯·馬什(Othniel Charles Marsh)所命名;另外,馬什同時也建立了新屬Camptonotus,後來改名為彎龍(Camptosaurus)。馬什當時與愛德華·德林克·科普(Edward Drinker Cope)進行一場長時間的競爭,名為化石戰爭。
馬什當時只敘述了模式標本的背部到尾巴的脊椎。馬什注意到脊椎的內部空腔,與細的脊椎壁,因此將它們命名為脆弱虛骨龍(Coelurus fragilis)。馬什認為虛骨龍的體型接近狼,食性應為肉食性。虛骨龍是莫里遜組所發現的第一個小型獸腳類恐龍,但馬什當時並不確定它們是否屬於恐龍。在1881年,他將虛骨龍歸類於恐龍,提出更多骨頭圖片,並建立虛骨龍目(Coeluria)與虛骨龍科,以包含虛骨龍屬。
虛骨龍的化石散布於挖掘地點,挖掘活動自1879年9月持續到1880年9月,馬什將部份化石建立為新種,敏捷虛骨龍(C. agilis),並從其中一對恥骨,研判敏捷虛骨龍的體型大於脆弱虛骨龍的三倍。在1888年,馬什將一個發現於馬里蘭州阿倫德爾組的趾爪,歸類於脆弱虛骨龍,該趾爪的年代是早白堊紀。儘管馬什與科普的競爭激烈,科普卻在1887年建立了兩個新種:鮑氏虛骨龍(C. bauri )與洛氏虛骨龍(C. longicollis),化石都發現於新墨西哥州的晚三疊紀地層。在兩年後,科普將這兩個種新建立為腔骨龍。
在1903年,亨利·費爾費爾德·奧斯本(Henry Fairfield Osborn)命名了嗜鳥龍,是莫里遜組的第二個發現的小型獸腳類恐龍。嗜鳥龍的化石是一個部份骨骸,發現於科莫崖北方的Bone Cabin採石場。在1920年代,查爾斯·懷特尼·吉爾摩爾(Charles W. Gilmore)在他的重要獸腳亞目研究中,提出虛骨龍與嗜鳥龍是同一種動物。這個分類法被其他科學家沿用,並持續了數十年。但是,沒有科學家將這兩種動物做正式的比較,也沒有人去統計哪些化石屬於虛骨龍。
吉爾摩爾曾提出脆弱虛骨龍、敏捷虛骨龍是同種動物,但直到80年代,約翰·奧斯特倫姆(John Ostrom)做了相關研究,才證實這個說法。這使脆弱虛骨龍的化石增加,而奧斯特倫姆也證實嗜鳥龍與虛骨龍是不同的物種。根據當時的不完整資料,戴爾·羅素(Dale Russell)曾提出敏捷虛骨龍是輕巧龍的一個種;奧斯特倫姆也指出其錯誤。奧斯特倫姆另外提出,馬什在1884年繪製的虛骨龍脊椎圖中,其中一個脊椎是另兩個脊椎拼合組成的;而剩下兩個脊椎中,有一個脊椎其實是發現於其他地點,屬於一個未命名的小型獸腳類恐龍。在1995年,懷俄明州發現的一個部份骨骸,一度被認為是個較大型的虛骨龍。數年後,這個新發現骨骸被建立為新屬長臂獵龍,是虛骨龍的近親。
虛骨龍目前只有一個有效種,模式種脆弱虛骨龍(C. fragilis),過去還有六個種被建立過。敏捷虛骨龍(C. agilis)由馬什在1884年建立,後來改歸類於脆弱虛骨龍。在1887年,科普根據在新墨西哥州發現的晚三疊紀化石,建立了鮑氏虛骨龍(C. bauri)與洛氏虛骨龍(C. longicollis);科普在1889年將這兩個種建立了新屬,腔骨龍。在1888年,理察·萊德克(Richard Lydekker)將哈利·絲萊(Harry Seeley)建立的達氏孔椎龍(T. daviesi)改為達氏虛骨龍(C. daviesi),該種化石發現於英格蘭的早白堊紀地層;後來獨立為新屬,鞘虛骨龍。纖細虛骨龍(C. gracilis)由馬什在1888年建立,化石是一些四肢骨頭,年代也是早白堊紀[。但吉爾摩爾在1920年代重新檢查這些骨頭時,只能鑑定出一個爪子。吉爾摩爾將其歸類於縴手龍,不屬於虛骨龍。最近的研究多認為纖細虛骨龍不屬於虛骨龍,是個疑名。但纖細虛骨龍不被認為屬於虛骨龍屬,也沒有成立個別的屬。嗜鳥龍過去曾被認為是虛骨龍的異名,其模式種被改為為郝氏虛骨龍(C. hermanni),直到奧斯特倫姆指出其錯誤。
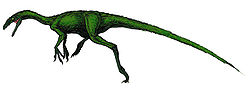
詳細介紹對於虛骨龍的知識,多來自於其中一副骨骸化石,這個化石發現於懷俄明州的科莫崖,包含眾多脊椎、部分骨盆、肩帶、以及四肢的大部分。但直到80年代,才將該骨骼完整的拼湊出來,而存放於皮巴第自然歷史博物館。另外,在猶他州的克利夫蘭勞埃德採石場發現的兩個前肢骨頭,有可能屬於虛骨龍。虛骨龍是種中小型恐龍,重量被估計為13到20公斤,身長約2.4米,臀部高度為0.7米。從重建的骨架模型來看,虛骨龍的脊椎骨長,而有長的頸部與身體,蹠骨長,因此後肢修長。顱骨小而長。前掌有3指,有銳利、彎曲的爪。
虛骨龍與人類的體型相比圖虛骨龍的頭部化石不多,只有挖掘地點所發現到的一個骨頭,可能是個部分下頜。該骨頭的保存狀況與外表,與虛骨龍的骨骸類似,但形狀較細長,可能不屬於該骨骼。這個骨頭的長度為7.9厘米,寬度為1.1.厘米。虛骨龍的脊椎長而低矮,神經棘短,脊椎壁細。頸椎有許多空腔,但分布不平均,大小不一。頸椎修長,長度是寬度的四倍,前後端有凹面。背椎較短,表面缺乏空洞,凹處較不明顯,形狀為沙漏狀。尾椎表面也缺乏空洞。
肩帶的唯一骨頭是個肩胛骨碎片。肱骨的側面是S狀彎曲,長度為11.9厘米,大於尺骨的9.6厘米。手腕有個半月形腕骨,類似恐爪龍,但跟人類的月骨(Lunatebone)不同。手指細而長。骨盆的唯一骨頭是兩個癒合的恥骨,尾端有明顯的柄部。股骨的正面呈S狀,長度約21厘米。蹠骨長,長度接近股骨。
共同發現虛骨龍、嗜鳥龍、長臂獵龍。
在莫里遜組,發現了三種外表相似的小型獸腳類恐龍,分別為:虛骨龍、嗜鳥龍、長臂獵龍,都屬於虛骨龍類,三個屬過去曾被互相混淆過。虛骨龍與嗜鳥龍已有較完整的敘述,可從許多生理特徵區別它們。例如:虛骨龍的頸部、背部較長,腿部與足部也較長。虛骨龍與長臂獵龍則較為類似,但也有許多不同點。例如:上臂與前臂及股骨的形狀、股骨的肌肉附著處、較長的背部脊椎、以及較長的蹠骨。
分類研究自從80年代以來,種系發生學研究逐漸增多,而虛骨龍通常被認為是虛骨龍類的分類不明屬,不屬於任何一個主要演化支。虛骨龍與美頜龍科、嗜鳥龍、原角鼻龍等屬,曾多次被歸類為基底虛骨龍類。在2007年,菲力·森特(PhilSenter)的長臂獵龍研究中,森特提出虛骨龍與長臂獵龍,是原始暴龍超科動物的近親。
虛骨龍有時被歸類於虛骨龍科,但虛骨龍科的物種卻不固定。在2003年,奧利佛·勞赫(OliverRauhut)提出虛骨龍科由虛骨龍、及美頜龍科的物種所構成。相反地,沒有科學家曾將虛骨龍歸類於美頜龍科過。在2007年,菲力·森特指出虛骨龍科只有虛骨龍、長臂獵龍兩個物種。
在採用種系發生學之前,虛骨龍科與虛骨龍下目都曾被當成“未分類物種集中地”,許多小型獸腳類恐龍被歸類於此,而且多為疑名。在80年代晚期的大眾恐龍書籍中,虛骨龍科包含超過10個物種,包括西北阿根廷龍科的福左輕鱷龍、偷蛋龍下目的小獵龍,而且虛骨龍科被當作是腔骨龍科的後代。在90年代初期的一些大眾讀物,虛骨龍科仍被當作“未分類物種集中地”。在這之後的物種數目就大幅縮減。
古生物學
虛骨龍發現於莫里遜組的第2與第5地層帶。莫里遜組是認為是個半乾旱的泛濫平原,有明顯的雨季與旱季。當地的植被十分多樣化,有河邊的走廊林、蕨類、樹蕨、針葉樹,以及疏林莽原。莫里遜組所發現的植物化石有:綠藻、真菌、苔蘚、木賊、蕨類、蘇鐵、銀杏、與數科針葉樹。動物化石則有:雙殼類、蝸牛、輻鰭魚類、青蛙、蠑螈、烏龜、喙頭蜥、蜥蜴、陸生與水生鱷形超目動物、數種翼龍目、大量的恐龍、以及早期哺乳類。
莫里遜組的恐龍包含:獸腳亞目的角鼻龍、異特龍、嗜鳥龍、蠻龍,蜥腳下目的迷惑龍、腕龍、圓頂龍、梁龍,鳥臀目的彎龍、橡樹龍、劍龍。早期哺乳類則有:柱齒獸目、多瘤齒獸目、對齒獸目、以及三尖齒獸目。
虛骨龍被認為是種小型肉食性恐龍,以昆蟲、哺乳類、蜥蜴等小型獵物為食。它們被認為是種行動快速的動物,比足部較短的嗜鳥龍還快。
化石群
四川達州市開江縣虛骨龍化石群
 川東恐龍化石集聚地
川東恐龍化石集聚地開江發現川東恐龍化石集聚地。這一重大發現,對於研究古生物的系統演化、生理特徵、生活環境以及川東地區地質構造等具有十分重要的科學價值。
2009年國慶期間,開江縣講治中學生物組師生到講治鎮伍家寨村雷公廟一帶山地實地採風,發現了很多貝殼化石和部分恐龍化石。發現這一情況後,該校立即向當地政府作了匯報。隨後,達州市文管所到現場進行考察核實發現,出土的化石中有二大塊恐龍脊椎化石,根據二大塊脊椎化石骨骼判定,恐龍長度應在20~30米左右,屬行動敏捷、兇猛的肉食性虛骨龍。
1964年夏,開江新寧鎮老三溝村發現了川東虛骨龍化石,現該化石完整地陳列在成都理工大學;1987年第二次全國文物普查時,曾在雷公廟、老三溝直線距離僅3公里左右的新寧鎮陡梯子、三里橋村,講治鎮雙飛村發現恐龍化石,如今再次發現的同屬虛骨龍化石。初步判斷該帶應屬川東虛骨龍的集聚地.
類群
 虛骨龍
虛骨龍按照現代新的分類觀點,虛骨龍類也已列入聯尾龍類,其中包括鳥的祖先。它在進化上的顯著特徵是:在齶部的外孔,後肢比前肢約長一半。它主要分布於亞洲及北美洲。目前,有許多不同的分類法將虛骨龍分成各種類群,這裡我們只介紹最重要的似鳥龍類和跑龍類。
似駝龍非常像現代的鴯鶓和鴕鳥。它們的頭較小,頸、尾巴和肢骨較長。似鴕龍生活在白堊紀晚期,因非常像鴕鳥而得名,由奧斯朋於1917年正式命名,它一般長約4米,也有2米多長的。它身體輕巧,有一個很小的頭,頜骨短而無力,口內無牙齒,有角質的喙嘴。它也有長的脖子和尾巴,以幫助平衡身體。長而苗條的後肢,比股骨長的脛骨,說明它跑得很快,主要靠輕捷纖長的後腿奔跑。它的腳很像鳥腳,腳上有三個腳趾,都有爪。前肢短,有三個手指,大拇指與其餘兩指對立,說明這是握物的器官。關於它的食性,由於這類恐龍發現於海岸旁,有人認為它可能是蹚水捕捉蝦和蟹為生的。奧斯朋則認為,它主要吃植物的嫩葉和幼芽。現在許多人認為它是雜食性的,既吃昆蟲和小動物如蜥蜴等,也可能偷吃恐龍蛋或剛生下來的小恐龍,一旦遇到大型的肉食性恐龍就馬上逃之夭夭,好在它跑得很快,一小時可達50公里。
虛骨龍類中的跑龍類,也是叫人特別感興趣的類群,它的代表就是恐爪龍。這種恐龍最早發現於美國蒙大拿州的中部,1969年由美國的奧斯特羅姆教授作了系統研究,出版了詳細的研究報告,揭開了它的奧秘。恐爪龍名稱的由來,是因為它生有恐怖的爪。它雖然只有三四米長,卻是恐龍家族中最兇猛的類群之一。它最大的特點是有結構特殊的腳。它的腳上有四個腳趾,趾上有利爪,但每個趾與爪的情況大不一樣。第一趾相當小,爪尖向後;第三趾與第四趾及爪的大小與粗細,也略有差別。爪較鈍,說明是有來行走的。很明顯,恐爪龍腳的細線在第三及第四趾之間通過,不像其他肉食性恐龍的腳軸線,通過第三趾。它用後肢行走,粗壯的踝關節和有力的肌肉使它在平地上行走時健步如飛。與其他肉食性恐龍相比,它的前肢已相對延長,前肢上有三個手指,指上也都有利爪,利爪相互對應,說明恐爪龍能用前肢握獵物。最吸引人的是恐爪龍的尾巴。它的尾巴絕大部分是棒狀長骨,這種棒狀骨是由部分特化的脊椎骨組成的,第一棒狀骨長度有八至十個脊椎骨的距離。這種僵硬的尾巴,可以用來保持身體平衡。它的頭也較大,眼眶骨大,說明它能“高瞻遠矚”地看到獵物。它的頜骨上布滿了利刃般帶鋸齒的牙齒,便於抓到獵物後大吃大嚼。由於在一次發掘時發現了五具恐龍爪的個體與一個軀體比單個恐爪大6倍的素食性恐龍,為此專家們認為,它是成群結隊地捕殺獵物的。它的獵物主要是一些素食性恐龍,當時的禽龍就是它的美餐。恐爪龍生活在白堊紀早期,除了北美洲外,蒙古也有發現。
在虛骨龍中還有著名的竊蛋龍。從它的名稱可以看出這是專門偷吃其他恐龍的動物。當美國的中央亞細亞考察團在蒙古首次發現竊蛋龍的骨架時,這個骨架正臥在原角龍的蛋上,於是研究這種恐龍的奧斯朋就認為它正在偷吃原角龍的蛋,所以叫竊蛋龍。這種恐龍體長2米,頭很小,在鼻骨的上方有骨質的隆起。有的竊蛋龍頭骨上有頂飾,有的則沒有,可能與雌雄有關。它的前肢上有在個手指,上面有尖銳彎曲的爪。拇指呈弧狀彎曲。指向其他兩指。這兩指比拇指大,這樣可以把獵物緊緊抓住。它的後肢也有三個腳趾,趾上也有利爪,腿粗壯有力,也是快跑的能手。它還有強壯的上下頜,具有很硬的喙嘴。1990年,中加恐龍計畫考察隊在內蒙古也發現了竊蛋龍的骨架,它正臥在一窩蛋上。據董枝明教授的看法,竊蛋龍能像鳥那樣孵蛋,所以看到這種場面,估計這條竊蛋龍可能是在孵蛋時遇到風暴而被風沙活埋的。現在許多恐龍專家認為它並不竊蛋,可能吃淡水中的蛤蜊或像鳥類一樣吃些大的漿果。它是雜食性恐龍。
似鳥龍也是虛骨龍類中的佼佼者。它生活在北美西部、蒙古以及我國的內蒙古。這種恐龍身長2.1~3.5米。它有一個相對說來較小的頭骨,但腦顱卻很大,說明它很可能有立體的視野。它的踝骨與脛骨癒合在一起,腳上有平直而窄的爪,在奔跑時能像跑鞋上的釘子那樣防止滑倒。這是恐龍世界的短距離快跑能手。它的腦子較大,腦顱大小與鴕鳥差不多,是恐龍家族中比較機警聰明的成員。它的尾巴很長,幾乎占了體長的一半,有的可達2米長。當它在崎嶇的坡地上奔跑時,這條長尾巴可以起平衡作用。它的牙齒小而多,下頜上就有三十五個,有人認為這是無所不吃的恐龍。它的眼睛比較大,在黃昏時刻也能捕捉小型的蜥蜴或哺乳類動物,甚至捕食昆蟲。
考古
 虛骨龍
虛骨龍《自然》雜誌發表中科院考古新成果
中國科學院古脊椎動物與古人類研究所徐星研究員及合作者在霸王龍類及虛骨龍類早期演化研究方向取得重要進展,有關成果發表於2月9日出版的英國《自然》雜誌上。鑒於該研究成果的重要性,《自然》雜誌還在同期配發了評論性文章。
最近幾十年的研究表明,鳥類起源於恐龍當中一支—虛骨龍類。這一假說的化石證據主要來自白堊紀,尤其是近年來發現於我國遼寧長羽毛的恐龍更是為這一假說提供了重要的證據。但由於已知最早的鳥類—發現於德國的始祖鳥生存於侏羅紀晚期,因此,有關鳥類起源更直接的證據應該存在於侏羅紀地層。在過去的6年中,徐星研究員與美國喬治華盛頓大學clark教授聯合領導的考察隊在我國新疆準噶爾盆地的戈壁地區進行了一系列的科學考察,此次報導的冠龍化石發現於2002年8月,是這一系列科考的成果之一。相關論文也是該項目發表於《自然》雜誌的第二篇論文。
冠龍化石產於沉積於大約160百萬年前的岩石中,遠早於已知霸王龍類,也是最早的虛骨龍化石代表之一。它的發現和研究為霸王龍類的早期演化以及虛骨龍類的系統發育提供了重要信息,再次證明體型變化等因素對於白堊紀虛骨龍類的形態產生了重要影響,干擾了虛骨龍類系統發育的復原;尤其是冠龍頭部嵴狀構造的發現指示了性選擇在生物進化當中的獨特作用。
這一論文由來自中國、美國和加拿大等3個國家的8名科研人員參與完成。有關研究工作得到了中國國家自然科學基金委、美國國家地理學會、美國國家科學基金委、中國科學院和美國喬治華盛頓大學的支持。
