
特徵
 花椰菜花葉病毒粒子的三維重組,示多層結構
花椰菜花葉病毒粒子的三維重組,示多層結構病毒粒子為等軸狀(直徑43~50 nm)或桿菌狀(30~35 nm × 60~400 nm),無包膜。花椰菜花葉病毒經低溫電鏡技術和計算機三維重構研究表明具有T=7的多層結構,病毒的外殼蛋白是糖基化的。花椰菜花葉病毒及其他球狀病毒屬的病毒具有相似的細胞病變特徵,在細胞質內形成內含體。
該科包括所有具雙鏈DNA的植物病毒。單分體基因組,開環狀dsDNA長約7.2~8.1 kb,DNA的一條鏈為編碼鏈,基因結構因屬而異,是分屬的主要特徵之一。當病毒進入細胞後,不連續的基因組連線成超螺旋DNA,並在細胞核中形成微染色體(minichromosomes),這些微染色體依靠寄主的依賴DNA的RNA聚合酶進行非對稱轉錄,轉錄一條比基因組長的轉錄本(35S或34S RNA),轉錄本有35~270 nt末端冗餘。此轉錄本即作為反轉錄成負鏈DNA的模板,又用於部分ORF的表達。花椰菜花葉病毒屬病毒產生一特定的mRNA(19S RNA)翻譯ORF 6,而碧冬茄脈明病毒屬、大豆褪綠斑駁病毒屬和木薯脈花葉病毒屬未報導產生亞基因組mRNA。水稻東格魯桿狀病毒屬的ORF 4由35S RNA剪下產生的RNA表達。與逆轉錄病毒相比,該科病毒的複製循環是游離的,不涉及整合階段,負鏈DNA的合成是由寄主細胞質中的tRNA(met)引發,而正負兩條鏈合成都是在病毒逆轉錄酶和核酶H的作用下進行的。位點特異的不連續性位於負鏈和正鏈DNA合成中的引物結合區,並在短距離上由新鏈取代舊鏈,而不連續形成一閉合環。各屬有2~8個ORF不等,所編碼產物一般為外殼蛋白、天冬氨酸蛋白酶、逆轉錄酶和RNA酶H。
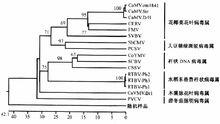 花椰菜花葉病毒科的系統發育樹
花椰菜花葉病毒科的系統發育樹病毒的自然寄主範圍較窄,碧冬茄脈明病毒屬、花椰菜花葉病毒屬、大豆褪綠斑駁病毒屬和木薯脈花葉病毒屬局限於侵染雙子葉植物,水稻東格魯桿狀病毒屬侵染單子葉植物,桿狀DNA病毒屬的寄主兩者均有。
花椰菜花葉病毒屬自然條件下由蚜蟲以半持久性方式傳播,也可以機械接種傳播。桿狀DNA病毒屬大多不能機械傳播,自然界中多數種由介體粉蚧以半持久方式傳播,水稻東格魯桿狀病毒屬由黑尾葉蟬傳播,需要有水稻東格魯球狀病毒的輔助。有些病毒分布範圍較窄,有些是世界性分布,尤其發生在溫帶地區。花椰菜花葉病毒屬等4個球狀病毒屬的免疫原性較強,2個桿菌狀病毒屬的免疫原性中等,各屬內的一些病毒之間有血清學關係。
分類
共6個屬。
| 中文名 | 學名 | 特徵 |
| 花椰菜花葉病毒屬 | Caulimovirus | 等軸粒子,7個ORF,蚜蟲傳播 |
| 碧冬茄脈明病毒屬 | Petunia vein learing-like viruses | 等軸粒子,2個ORF,無介體 |
| 大豆褪綠斑駁病毒屬 | Soybean chlorotic mottle-like viruses | 等軸粒子,8個ORF,無介體 |
| 木薯脈花葉病毒屬 | Cassava vein mosaic-like viruses | 等軸粒子,5個ORF,無介體 |
| 桿狀DNA病毒屬 | Badnavirus | 桿菌狀粒子,3個ORF,粉蚧傳播 |
| 水稻東格魯桿狀病毒屬 | Rice tungro bacillifrom-like viruses | 桿菌狀粒子,4個ORF,黑尾葉蟬傳播 |
