
生物分布
正文
即生物的地理分布。任何生物分類單位(種、屬、科或種以下的亞種、變種等)在地球表面分布的範圍叫做它的分布區,不同分類單位的分布區大小和形狀差別很大,這既取決於生物本身的散布能力,也取決於它所遇到的外界條件;此外,還決定於歷史因素等。生物的散布能力是由它的繁殖速度和它的散布方式決定的。凡產生種子或孢子多、繁殖快的,它的散布能力就大,低等植物往往產生驚人數量的繁殖體,它們的散布能力就比產生較少繁殖體的種子植物大得多。動物也是一樣,每窩仔僅數個的哺乳動物(靈長目、鯨目等),其散布能力遠遠不如大量產卵的昆蟲。風布和動物布與不利用外界能源的散布(重力散布、營養繁殖)方式相比,能到達的距離更遠。對於少數生物,水布也是遠距離散布的一種方式。在交通發達的現代,人類幫助某些生物的散布,其速度之快,所達距離之遠,超過其他動力。
 生物分布
生物分布限制生物分布的因素 生物的本性是產生儘可能多的後代,占領儘可能大的地盤(分布區),然而由於外界條件的限制,它並不能實現這一本性。限制生物分布的外界因素來自兩方面:環境影響;同其他生物的相互作用。
環境影響 生物只能生存於一定的環境條件範圍內。一般說來,氣候和土壤是決定植物分布的主要環境條件;大多數動物以植物為食或以其為蔽所,因此動物的分布往往與植物的分布相合。從赤道到極地和從沿海到內陸的氣候變化不僅伴隨著植物分布的改變,而且也伴隨著動物分布的改變。
有時分布區的外形不決定於氣候,而決定於基質。例如鹽生植物的分布與鹽漬土的分布相一致。
環境影響有時表現為明顯的“地理障礙”。例如,海洋對於絕大多數陸生生物是不可逾越的障礙,陸地對於水生生物也是一樣,所以許多生物的分布區以岸線為界。蘇伊士運河未鑿通前,地中海和紅海具有極不相同的動物區系,運河挖通後,某些紅海種的分布區延伸到地中海東部。
地形對於陸地棲居者來說也是重要的分布障礙,連續不斷的山鏈(例如喜馬拉雅山、秦嶺、天山、祁連山)對於生活在低海拔地區的種有如一堵不可通過的牆,因此山兩邊可以發現很多不同的種類。大型湖泊和河流有時也成為陸地棲居者通過的障礙。
同其他生物的相互作用 一個生物種的分布可能決定於同其他種的相互作用。例如有毒植物牛心朴子目前在鄂爾多斯一帶廣泛分布,這不僅由於它本身不為牲畜所吃,還由於該地區長期強度放牧,減少了其他植物對它的競爭壓力;各類針茅數量的日漸減少則是牲畜直接大量啃食所造成的。
一定植物群落常常成為某些動物分布的限制,特別是占領廣闊空間的群落,這種限制作用尤其顯著。例如,對於大多數熱帶森林動物來說,稀樹草原是它們分布的障礙,但同時熱帶森林也是稀樹草原棲居者分布的障礙。在這種情況下,障礙作用既是環境條件的改變,也是競爭的結果。
生物的局部分布 從小範圍看,生物個體分布在不同種之間差別很大,一般有3種基本類型(圖1):
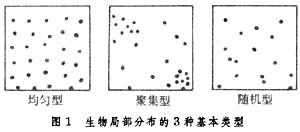 生物分布
生物分布聚集分布 個體成群或聚集分布。聚集型形成的原因有幾種:或者由於個體相互接近有利(如動物群);或者由於環境的不均勻性,有利於生物在某一部分存在而不利於在另一部分存在;或者由於繁殖的特性造成(植物的匍匐莖或根莖的無性繁殖)。大多數動物趨向於聚集型分布。土壤或森林枯枝落葉層中大多數節肢動物都成群或聚集分布。
隨機分布 個體的分布是偶然性的。在特別一致的環境裡,生物自由選擇定居地點,這有利於隨機型分布的出現。例如在潮帶間的泥灘,某些蛤類通常呈隨機型分布。
生物的大範圍分布 由於起源、歷史、生態要求和環境條件的差異,不同物種分布區的大小和外形極其多種多樣,但幾乎所有物種都在地球表面占據不規則形狀的面積。分布區形狀的不規則性在大多數情況下由環境特點所決定。
世界性分布區 指覆蓋地球大部分面積的分布區。目和科的世界性分布較多。動物中的輪蟲類,種子植物中的菊科、禾本科、豆科和莎草科都具有世界性分布。屬和種只有很少是相對世界性的。大約有20種左右種子植物可以歸入世界性分布,它們包括許多水生和沼澤植物,如浮萍、蘆葦、香蒲、水燭等。伴隨人類散布的雜草,有些也是世界性的,如早熟禾、薺菜、蒲公英、大車前等。動物中的家蠅和褐家鼠等的分布也遍及全世界。
環大陸和環海洋分布區指占據某一緯度範圍的陸地或海洋的分布區。包括環北極分布,如拉伯蘭苔草;北半球中緯環陸分布,如驢蹄草,茶藨子;泛熱帶分布,如棕櫚科植物(圖2)、含羞草、馬錢子、羊蹄甲等;南半球高緯環陸分布,這是斷續分布,以某些古老類群的生物為特徵,如原始導管植物,有爪綱以及Styloniceae科的等足目甲殼類。
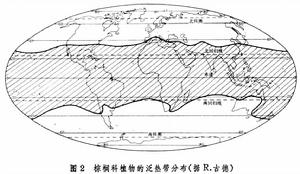 生物分布
生物分布特有分布區 與世界性分布相反,特有分布只限分布於某一有限地域。特有分布區的面積可能多種多樣,目、科、屬的特有分布區面積一般較大,例如澳大利亞、塔斯馬尼亞和紐幾內亞總起來形成單孔目的特有地域。對於種或變種來說,其特有分布區可能面積很小。例如就現在所知,金佛山蘭(Tangtsinia nanchuanica)只產於中國四川省南川縣的金佛山。
從進化觀點看,特有現象可能是兩種完全不同的進化過程的結果:①古特有種,或稱殘遺特有種,都是古老的物種,早先的分布區極為廣闊,後來由於氣候改變或由於適應力較強的其他種的競爭,其分布區逐漸縮小。所以古特有種大多處於消失的過程中。例如銀杏,在中生代三疊紀和侏羅紀時遍布全世界,在第四紀冰川期,中歐和北美的銀杏都已絕滅,只在中國浙江天目山一帶還保存有野生種。所以銀杏目前的分布區(野生)很小,是古特有分布區。水杉(Metasequoia glyptostroboides)在第三紀時也廣布於北半球,現在野生狀態的個體僅在中國湖北西部利川縣和四川東部萬縣的少數幾個地區能夠見到,也是古特有種。被隔離的山鏈往往是特有種集中的地方。例如在非洲撒哈拉山區的阿哈加爾山脈,特有種占植物的40%。②新特有種,或稱年幼特有種,是一類不久以前在局部地區產生的特有種。由於形成時間不久,往往只達到亞種級。例如生活於不列顛和中歐湖泊中的一種淡水鮭魚鴉巴沙(Coregonus),在每一湖泊中都有不同形態的個體。在不久以前才與歐洲大陸分離的不列顛群島上,陸生動物中新特有種數目特別多。
替代分布區 從相鄰兩個區域的同樣生境起源,或從同一區域的兩個不同生境起源的兩個近緣分類單位相互取代的現象叫替代現象,它們的分布區是替代分布區。前一種情況是地理替代;後一種情況是生態替代。前者如地中海流域的油橄欖(Olea europaea),它的野生形式在撒哈拉山區為非常接近的O.laperrinei所替代;後者如中國南方的狗脊 (Woodwardia japonica)和鐮狗脊W.unigemmata),狗脊生長於酸性土上,而鐮狗脊則要求石灰性土。在砂質岩(或第四紀紅層)和石灰岩相間分布的山坡,就可以看到狗脊和鐮狗脊相間出現。巴拿馬地峽東岸和西岸動物區系的相互替代是海洋替代的極好例子。地峽大約在上新世開始形成,使兩岸相互隔開。目前許多魚和軟體動物可以相近形式成對出現,其中每一對中的一個出現在大西洋近岸水域,另一個就出現在太平洋近岸水域。
間斷分布區 或稱不連續分布區。有些分類單位的分布區斷裂成幾個部分,並且彼此相距很遠。相似的或有密切關係的甚至相同的種出現在地球的不同部分,形成相互隔離的分布範圍,這樣的分布區叫間斷分布區或不連續分布區。間斷分布區有多種多樣的類型。例如,著名的第三紀殘遺植物鵝掌楸屬中,一種(即鵝掌楸)分布於中國南方,另一種(北美鵝掌楸)分布於北美東部,為洲際間斷(圖3)。不僅鵝掌楸屬,整個木蘭族也是洲際間斷。
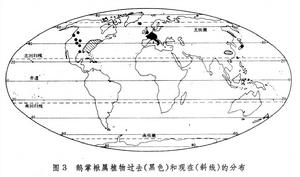 生物分布
生物分布特別令人驚奇的是兩極間斷分布(或稱雙極分布)。同一個屬的不同種,甚至同種,既出現在北極附近,也出現在南極附近,而中間地區卻沒有,例如岩高蘭屬主要具有環北極分布區,但又出現在南美的南端。海洋動物中也有雙極分布的例子,例如北極海參(Psolus)和南極海參。
關於造成間斷分布的原因曾有許多爭論。一種解釋是遠距離散布,特別是往返於南北半球之間的候鳥的遷徙,可能是傳布繁殖體的重要媒介。候鳥遷徙對解釋某些植物的兩極間斷分布有幫助,但不能解釋有些洲際間斷,如上面談到的鵝掌楸在東亞和北美東部之間的間斷。為什麼同樣緯度、氣候條件相似的北美西部沒有鵝掌楸,這難以用候鳥遷徙解釋。有些人提出相似種在地表不同的和遠離的部分完全獨立地起源的假說,但很少證據能夠證明這點。普遍接受的假說是:相似的或遺傳上有關係的生物類型在過去必然來源於一共同祖先,從共同的起源中心向外侵移到它們所占據的地區,而目前分布區的不連續性是以後發生的環境改變──“障礙”造成的,這種障礙把它們目前占領的地區分開。例如,北極-高山間斷分布是由於在冰川期間北半球的冰蓋向南延伸到中緯度,覆蓋了歐亞大陸和北美的廣大地
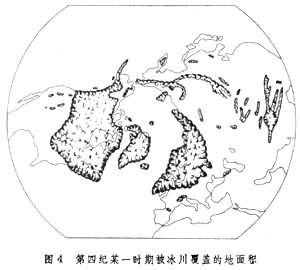 生物分布
生物分布有些間斷分布很難從生物的繁殖能力、散布方式和生境特點等方面圓滿解釋,必須考慮歷史因素。例如假山毛櫸屬(Nothofagus)目前的分布範圍是南半球的智利南端、紐西蘭、澳大利亞、新喀里多尼亞和紐幾內亞,但非洲沒有。有人研究了這個屬的散布特徵得出結論:它的種子不適於跳躍式散布,因而不能越過廣闊的鹹水海洋,它現在的不連續分布是岡瓦納古陸破裂產生的結果。A.L.韋格納的大陸漂移學說,隨著新近板塊理論的出現已得到普遍承認。按照大陸漂移說,大陸曾經是連線在一起的,後來逐漸裂開。在中生代的某個時候,全部世界的陸地組成兩塊大的大陸:勞亞古陸和岡瓦納古陸。勞亞古陸由現在的北美和歐亞大陸組成;岡瓦納古陸由現在的南極大陸、澳大利亞、南美、非洲和印度組成。這兩塊古陸中間隔著古地中海。後來勞亞古陸和岡瓦納古陸破裂成較小的部分。岡瓦納古陸的碎塊向外漂移開,就形成現在南半球的幾塊陸地。假山毛櫸是岡瓦納屬,它的分布限於岡瓦納古陸,雖然現在在南極大陸沒有假山毛櫸的分布,但南極大陸上有兩個地點發現了假山毛櫸的花粉化石,所以該屬今天的間斷分布應是岡瓦納古陸破裂的結果形成的。
地球的生物地理分區 如果把地球不同部分的植物和動物種類進行對比,就會發現彼此有明顯的差別,兩地相距越遠差別越大。雖然可能有些分類單位彼此共有,但通常是許多分類單位只出現在某一地區,而不出現在另一地區。這就有可能把地球劃分為不同的植物地理區域(見彩圖)和動物地理區域(見彩圖),或者是同時考慮植物和動物的生物地理區。無論植物、動物或生物地理區域都是客觀的存在,都是科學研究的實體。
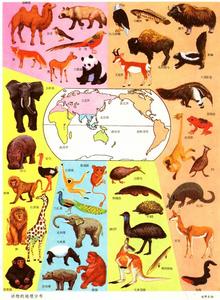 生物分布
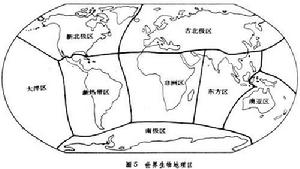
生物分布E.C.皮耶盧1979年對動物和植物給予同樣的強調。把施密特的省結合成 8個“生物地理區”,即古北極區、新北極區、新熱帶區、非洲區、東方區、澳亞區、南極區,此外還包括一個大洋區(圖5)。
 生物分布
生物分布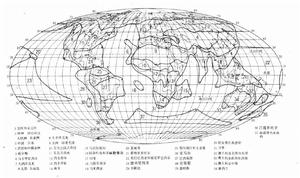 生物分布
生物分布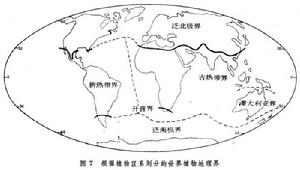 生物分布
生物分布動物地理區域的劃分,從A.R.華萊士以來公認的方案也是將地球陸地分為6個動物地理界,每一個或多或少包括一個主要大陸,並且互相間被海洋或山脈分開。它們是:古北界、新北界、新熱界、非洲界、東洋界和澳洲界。
較新的一個動物地理劃分方案是K.P.施密特1954年提出的,他區分出4個不同等級的單位:界、區、亞區和省。他一共劃分出3個界(北界、新界、南界)、5個區(古熱區、全北區、新熱區、澳大利亞區、大洋區)、13個亞區和32個省。
海洋可以分為兩個界:大陸架和深海。200米深等深線是大陸架和深海的分界線。這是光透入的下限。在此限度以上的海洋生物能進行光合作用。深海從來沒有成為過陸地,那裡動物區系分布簡單,在世界範圍內比較一致,具有廣泛分布的世界性科。而大陸架實際是大陸的一部分,沉沒在陸緣海之下,動植物區系較為複雜多樣。
