
機制
與配體結合
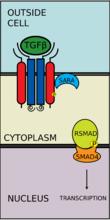
TGF-β超家族包括:骨形成蛋白(BMP),生長分化因子(GDF),抗繆勒式管激素(AMH),激活素(Activin),Nodal及TGFβ。信號轉導開始時,TGFB超家族配體與TGF-βII型受體結合。II型受體是一種絲氨酸/蘇氨酸激酶受體,它催化了I型受體的磷酸化。每種配體與一種特定的II型受體相結合。在哺乳動物中,現有七種已知的I型受體和五種已知的II型受體。
有三種激活素:激活素A,激活素B,激活素AB。激活素參與胚胎髮生和骨發生。它們也調控包括垂體激素、性激素、下丘腦激素及胰島素在內的多種激素。它們還是神經細胞存活因子。
骨形成蛋白(BMP)與2型骨形成蛋白受體(BMPR2)結合。它們參與包括骨形成、細胞分化、前後軸分化、生長及動態平衡在內的等多種細胞功能。
TGF-β超家族包括:TGFβ1, TGFβ2, TGFβ3。就像BMP一樣,TGF-β參與胚胎髮生和細胞分化,但它們還參與細胞凋亡等其它功能。它們與2型TGF-β受體(TGFBR2)結合。
Nodal與IIB型激活素受體(ACVR2B)結合。之後它或者與IB型激活素受體(ACVR1B)結合形成受體複合物,或者與IC型激活素受體(ACVR1C)結合。
受體-配體結合反應在局部發生時,該類反應被歸為旁分泌信號轉導。
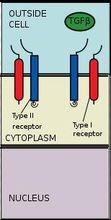 TGFβ配體與受體結合
TGFβ配體與受體結合 受體招募與磷酸化
TGF-β配體與一個II型受體二聚體結合,該II型受體二聚體招募一個I型受體二聚體,並與配體形成異源四聚體複合物。這些受體是絲氨酸/蘇氨酸激酶受體。它們有一個富含半胱氨酸的胞外域,一個跨膜域和一個富含絲氨酸/蘇氨酸的胞內域。I型受體的GS域由一系列大約三十個絲氨酸-甘氨酸重複組成。TGF-β家族配體的結合造成受體翻轉,這樣它們的胞內激酶域就處於一個易於發生催化反應的方向。II型受體將I型受體的絲氨酸殘基磷酸化,I型受體再激活蛋白。
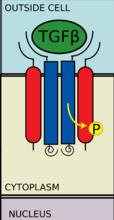 II型受體招募I型受體並將其磷酸化
II型受體招募I型受體並將其磷酸化 SMAD 磷酸化
有五種受到受體調控的SMAD蛋白:SMAD1, SMAD2, SMAD3, SMAD5及SMAD9(有時也被記作SMAD8)。主要有兩種胞內通路與這些R-SMAD蛋白相關。TGF-β、激活素、Nodal以及一些生長分化因子(GDF)被SMAD2及SMAD3調控,而骨形成蛋白(BMP)、抗繆勒式管激素(AMH)和個別生長分化因子(GDF)被SMAD1、SMAD5及SMAD9調控。R-SMAD蛋白與I型受體的結合由含蛋白質的FYVE鋅雙指結構域(zinc double finger FYVE domain)調控。SARA(The SMAD anchor for receptor activation,受體活化時SMAD的錨定點)和HGS(Hepatocyte growth factor-regulated tyrosine kinase substrate,受肝細胞生長因子調控的酪氨酸激酶的底物)是兩種調控TGF-β通路的蛋白。
SARA存在於初級內體中,這些初級內體通過內吞作用攝入受體複合物,這個過程受到格線蛋白調控。SARA招募R-SMAD蛋白,SARA使R-SMAD蛋白能與I型受體的L45域結合。SARA調整R-SMAD的方向,使得其羧基端上的絲氨酸殘基朝向I型受體的催化區域。I型受體將R-SMAD的絲氨酸殘基磷酸化,從而導致R-SMAD中的MH2結構域發生形態改變,進而與受體複合體及SARA脫離。
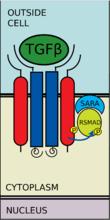 I型受體將R-SMAD磷酸化
I型受體將R-SMAD磷酸化 與CoSMAD結合
磷酸化的RSMAD與coSMAD(如SMAD4)有很強的親和力,並與coSMAD形成複合體。在這個反應中,RSMAD的磷酸基並沒有作為coSMAD的停泊位點發揮作用,而是磷酸化打開了一段胺基酸,從而RSMAD和coSMAD能相互反應。
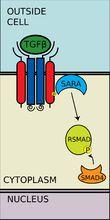 R-SMAD與coSMAD結合
R-SMAD與coSMAD結合 轉錄
磷酸化的RSMAD/coSMAD複合物進入細胞核,與轉錄啟動子及轉錄輔助因子結合,引起DNA轉錄。
成骨蛋白引起參與骨發生、神經形成及腹部中胚層分化的mRNA的轉錄。
TGF-β引起參與細胞凋亡、細胞外基質再生及免疫抑制的mRNA的轉錄。它也與細胞周期中的G1期阻滯有關。
激活素引起參與性腺生長、胚胎分化及胎盤形成的mRNA的轉錄。
Nodal引起參與左右軸分化、中胚層和內胚層形成的mRNA的轉錄。
 R-SMAD-coSMAD複合體進入細胞核
R-SMAD-coSMAD複合體進入細胞核 通路調控
TGF-β信號通路參與許多細胞過程,因此受到頻繁的調控。TGF-β信號通路有多種正反饋和負反饋調節機制,如配體和R-SMAD的激動劑,誘餌受體,R-SMAD和受體被泛素化等。
配體激動劑/拮抗劑
脊索蛋白和頭蛋白都是骨形成蛋白(BMP)的拮抗劑。它們與BMP結合,阻礙其與受體的結合。有研究顯示,脊索蛋白和頭蛋白使中胚層形成背部組織/器官,二者都在非洲爪蟾的背唇中被發現,另外還將特定的上皮組織轉化為神經組織。頭蛋白在軟骨形成和骨骼形成中起關鍵作用。
DAN家族的蛋白也與TGF-β家族成員拮抗。這些蛋白包括Cerberus,DAN以及Gremlin。這些蛋白包含可以形成二硫鍵的九個保守半胱氨酸。一般認為DAN與GDF5,GDF6及GDF7拮抗。
卵泡抑素與激活素結合併抑制激活素,它直接影響促卵泡激素的分泌。卵泡抑素也與前列腺癌密切相關,其基因的突變可能阻礙它對激活素的作用,而這種作用具有抗增殖的性質。
Lefty蛋白是TGF-β的一種調控因子,它參與了胚胎形成中的軸形成。它也是TGF超家族的成員。在鼠類胚胎中,這種蛋白在左側不對稱地表達,因而與左右分化相關。Lefty通過抑制R-SMAD的磷酸化產生作用,這樣的過程通過一種持續性活化的TGF-βI型受體及Lefty活化的下游過程實現。
也有研究發現基於藥物的拮抗劑,例如SB431542,它選擇性地抑制ALK4, ALK5, and ALK7。
受體調控
轉化生長因子受體3(TGFBR3)是發現的數量最多的TGF-β受體,它沒有已知的信號域,但它能通過結合TGF-β並將其呈遞給TGFBR2來增強TGF-β配體與TGF-βII型受體的結合。TGF-β信號通路的下游靶標之一GIPC與TGFBR3的PDZ結構域結合,抑制其蛋白酶體降解,進而增強TGF-β活性。它可能也是激活素RIL的抑制素複合受體。
BAMBI(BMP and activin membrane bound inhibitor,BMP和激活素膜結合抑制劑)與I型受體有相似的胞外域,它缺少一個絲氨酸/蘇氨酸蛋白激酶胞內域,因此它是一個虛擬受體。BAMBI與I型受體結合,阻止其被激活。它是TGF-β信號通路中的負調控因子,可能在胚胎形成中抑制TGF-β表達。它的表達需要BMP信號。
FKBP12與I型受體的GS域結合,抑制II型受體將I型受體磷酸化。一般認為FKBP12及其同系物在沒有配體時抑制I型受體活性,因為配體結合後它會斷開與I型受體的連線。
R-SMAD調控
抑制性SMAD的功能
SMAD家族中還有兩種抑制性蛋白(I-SMAD),即SMAD6和SMAD7。它們參與負反饋調節,並在TGF-β信號通路的調控中起關鍵作用。與其它SMAD蛋白一樣,它們有一個MH1結構域和一個MH2結構域。SMAD7與其它R-SMAD蛋白競爭,與I型受體結合,並抑制其磷酸化。它們儲存在細胞核中,一旦TGF-β被激活就進入細胞質並結合I型受體。SMAD6與SMAD4結合,阻礙其它R-SMAD蛋白與coSMAD蛋白結合。在TGF-β信號通路中,I-SMAD的表達水平提高,這意味著它是TGF-β信號通路的下游靶標。
R-SMAD的泛素化
E3泛素蛋白連線酶SMURF1和SMURF2調控SMAD的表達水平。與酶結合的E在酶上將泛素傳遞給RSMAD,導致其泛素化及蛋白酶體降解。SMURF1與SMAD1及SMAD5結合,而SMURF2 與SMAD1, SMAD2, SMAD3, SMAD6及SMAD7結合。SMURF2增強了SMAD7的抑制活動,而減少了SMAD2的轉錄活動。
一覽表
| TGF-β超家族配體 | II型受體 | I型受體 | R-SMAD | coSMAD | 配體抑制劑 |
| 激活素A | ACVR2A | ACVR1B(ALK4) | SMAD2,SMAD3 | SMAD4 | 卵泡抑素 |
| 生長分化因子1 | ACVR2A | ACVR1B(ALK4) | SMAD2,SMAD3 | SMAD4 | |
| 生長分化因子11 | ACVR2B | ACVR1B(ALK4),TGFβRI(ALK5) | SMAD2,SMAD3 | SMAD4 | |
| 骨形成蛋白 | BMPR2 | BMPR1A(ALK3),BMPR1B(ALK6) | SMAD1,SMAD5,SMAD8 | SMAD4 | 頭蛋白,脊索蛋白,DAN |
| Nodal | ACVR2B | ACVR1B(ALK4),ACVR1C(ALK7) | SMAD2,SMAD3 | SMAD4 | Lefty |
| TGD-β | TGFβRII | TGFβRI(ALK5 | SMAD2,SMAD3 | SMAD4 | LTBP1,THBS1,核心蛋白聚糖 |
