
概述
 蓖麻
蓖麻蓖麻毒蛋白(Ricin)是蓖麻種子中的一種毒蛋白,含量占籽重的1%-5%。在1887年,Dixson首次證明了蓖麻籽的毒性是由一種蛋白引起的。19世紀60年代,Stillmark詳細地研究了這種毒素蛋白,並建議命名為“Ricin”。此後,蓖麻毒蛋白所具有的特殊的理化性質和生物學特性引起許多生物化學家的濃厚興趣和關注。研究表明,蓖麻毒蛋白對所有哺乳動物真核細胞都有毒害作用,而對某些惡性腫瘤細胞毒性更強。這使它在醫學上成為用於殺傷腫瘤細胞的首選毒素之一。Ricin是天然藥物中最毒的蛋白之一,其小鼠LD50為7-10μg/kg,成人致死量為7mg,平均一分子Ricin就足以殺死一個細胞。
蓖麻毒蛋白由A,B兩條鏈構成,具有很強的細胞毒性,一旦進入胞質溶膠,毒蛋白A鏈便會催化核糖體的脫嘌呤作用,1min便可導致1500個核糖體失活。蓖麻毒蛋白在醫學上廣泛用作抗癌免疫毒素。中國是蓖麻的主要生產國,蓖麻的研究處於世界領先地位。隨著蓖麻油需求量的增大,大量的蓖麻脫脂粕產生,但由於含有毒性物質,而限制了蓖麻的綜合利用。
物化性質
生化組成
 蓖麻毒蛋白
蓖麻毒蛋白蓖麻毒蛋白的胺基酸組成(質量百分比)如下:賴氨酸-1.5%,擷氨酸-2.9%,甘氨酸-2.0%,異亮氨酸-3.6%,組氨酸-0.9%,色氨酸-0.8%,亮氨酸-3.8%,苯丙氨酸-2.3%,蘇氨酸-2.%,膚氨酸-1.6%,天冬氨酸-10.3%,蛋氨酸-0.9%,谷氨酸-6.8%,精氨酸-12.7%,絲氨酸-8.2%。由此可見蓖麻毒蛋白中精氨酸、天冬氨酸、絲氨酸和谷氨酸的含量較多。而A鏈色氨酸200、精氨酸180、谷氨酸177、酪氨酸123和酪氨酸80在胺基酸序列中是幾個保守的非極性胺基酸,它們對穩定活性中心起了一定的作用。由於A鏈的賴氨酸的含量低,所以可防止泛素化和泛素介導的蛋白酶水解,有研究證明賴氨酸被移除不會影響A鏈的活性、結構、穩定性,如果在上面附加4個賴氨酸殘基,A鏈的降解速度會明顯加快。
蓖麻毒蛋白為白色粉末或結晶型固體,無味,不溶於乙醇、乙醚、氯仿、甲苯等有機溶劑,溶於稀酸或鹽類水溶液,在飽和的硫酸銨溶液中能沉澱析出。在沸水中或加壓蒸汽處理可使Ricin凝固變性,失去毒性,但在乾熱的情況下變性很小。蓖麻毒蛋白在蓖麻籽中含量為1%~5%,也有含量為0.5%~15%的報導,其在蓖麻的根、莖、葉中也有一定含量。熱榨油形成的蓖麻粕中Ricin活性趨近於零,而在冷榨油形成的蓖麻粕中Ricin活性較高,因此Ricin在蓖麻粕中活性取決於榨油方式。與一般蛋白質相比,Ricin對熱、酸、鹼比較穩定,在半乳糖溶液中可以保存數月而不失活。
分子鏈結構
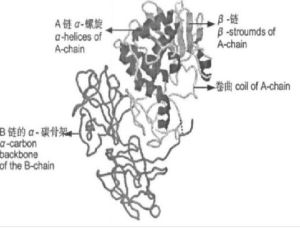
Ricin由兩個肽鏈以二硫鍵共價相連線,作為糖蛋白,Ricin含有共價結合的糖分子,糖的主要組成是甘露糖、葡萄糖和半乳糖。蓖麻毒蛋白的一級結構分析已由Funatsu等人完成。兩條多膚鏈分別稱為A鏈(RicinchainA,RTA)和B鏈(RicinchainB,RTB)。RTA是活性鏈,也是是毒性鏈,是一種糖苷酶,相對分子質量約為31000。RTB是結合鏈,有凝集素的活性,相對分子質量約為34000,B鏈上含有兩個半乳糖結合位點,能與細胞上含半乳糖的糖蛋白或糖脂結合。兩者間由二硫鍵連線。RTA和RTB都有糖基化的側鏈。蓖麻毒素的一級結構分析己由Funats等完成。結果顯示:A鏈含有約263個胺基酸殘基,第10個殘基Asn為糖基化部位,接有(G1cNAc)2(Man)4寡糖鏈。RTA是由267個胺基酸組成的球形蛋白,含有8個α螺鏇和8個β轉角和一些無規則捲曲等構象單位,活性中心為Arg180;RTB是由262個胺基酸組成的啞鈴形蛋白。通過生化和突變分析,證明RTB至少有3個半乳糖結合位點,可與細胞膜上的糖,如半乳糖和N-乙醯多巴胺形成氫鍵。RTA的Cys259和RTB的Cys4由二硫鍵結合,藉助RTB的攜帶作用而使RTA進入細胞發揮其毒性。不同蓖麻及其變種的毒蛋白,其胺基酸序列不完全相同。Ramesh等分離出2種不同的毒蛋白RicinD和RicinE,而Cawley等分離出3種不同的毒蛋白Ricin1、Ricin2和Ricin3。這些蛋白的等電點從5.9到8.8不等。A鏈中只有兩個Lys殘基,一個位於N端第4位,一個位於C端附近,對於A鏈的毒性作用極為重要。B鏈是由260個胺基酸殘基組成,並有4個分子內的二硫鏈,有兩條寡糖鏈(G1cNAc)2(Man)6和(G1cNAc)2(Man)7,分別接在第93位和133位Asn殘基上,故分子量較A鏈高。B鏈兩條寡糖鏈末端甘露糖殘基可以和網狀內皮細胞特別是巨噬細胞結合,後者細胞表面富含甘露糖受體,可優先攝取蓖麻毒素,這對於毒素髮揮生物功能有重要的作用。其A鏈(RTA)在B鏈(RTB)的協助下,容易穿過細胞膜,破壞核蛋白體60S亞單位,抑制蛋白質的合成,導致細胞死亡。Ricin的一級結構表明,A鏈由263個胺基酸殘基組成,分子量32000,是活性鏈,第10個殘基Asn(天門冬醯胺)已糖基化,接有(GlcNac)2:(Man)4寡糖鏈。A鏈中只有兩個Lys殘基,一個位於N端第四位,一個位於C端附近,它們對A鏈的毒性作用至關重要。B鏈是由259個胺基酸殘基組成的序列,含有兩個寡糖鏈(GlcNac)2(Man)8和(GlcNac)2(Man)7,分別與第93位和第133位上兩個Asn殘基相連,分子量34000。不同品種的蓖麻及其變種的毒蛋白的胺基酸序列不完全相同。A鏈(RTA)和B鏈(RTB)都有糖基化側鏈,兩者間由二硫鍵連線RTA和RTB均已克隆並在大腸桿菌中表達。A鏈共分為三個結構域。結構域1為N端117個胺基酸殘基,約占整個鏈長的40%,由6個β摺疊和2個α螺鏇組成.結構域2為118-210位也由近40%胺基酸殘基組成,主要由5個α螺鏇組成,其中E螺鏇最長,由20個胺基酸殘基組成(161-180位),長度超過5個螺鏇,位於整個分子中心,並有一個偏向C端約30度彎曲,摺椅彎曲使谷氨酸177,精氨酸180伸向分子表面形成活性中心。剩下的20%殘基為結構域3,主要由無規則捲曲組成,靠近C端富含疏水胺基酸,它可能對蓖麻毒蛋白A鏈的跨膜運輸起重要作用,結構域3一側形成活性中心的一部分,另一側通過疏水相互作用與B鏈結合。
天然A鏈是一種高度糖基化的蛋白,容易被肝細胞識別並加以清除,用原核系統表達重組的蓖麻毒蛋白A鏈(recombinantRicintoxinAchainrRTA)不但可以排除B鏈的影響,而且也不發生糖基化,彌補了天然A鏈的缺陷。1992年美國芝加哥大學Morris等人研究得出:在蓖麻毒蛋白267個胺基酸殘基中,222個(約83%)可逐個刪除而不影響A鏈識別和催化活性,但允許刪除片段都較短,僅一個片段含20個胺基酸,其餘是5個、2個胺基酸小片段。蛋白內部一些疏水性殘基,還有部分α-螺鏇和β-摺疊中殘基也可刪除,這使A鏈有了相當強的彈性,它可彌補結構上一定程度的變化,從而保持其生物活性。B鏈是結合鏈,分子量約為34000,由259個胺基酸殘基組成,空間結構似啞鈴,每一邊都有一個半乳糖結合位點。B鏈由兩個結構域組成,這兩個結構域有很高的同源性。1-135位胺基酸組成結構域1,136-262位胺基酸組成結構域2,從結構上看這兩部分的胺基酸序列有32%相同,每一結構域又可分為α,β,γ亞結構域以及λ連線肽,亞結構之間相互作用形成疏水核,來穩定B鏈的二維構象,結構域1的α和結構域2的γ亞結構域有糖結合活性,因此推測B鏈由一個具有結合半乳糖性質的多肽,通過基因重複與融合而形成,與獨立進化的蓖麻毒蛋白A鏈基因融合而形成蓖麻毒蛋白。蓖麻毒蛋白B鏈與糖的結合主要通過氫鍵,這也決定了對糖構型的專一性,同時蓖麻毒蛋白B鏈與糖相互作用的位點比較少,這也與蓖麻毒蛋白B鏈與半乳糖結合能力低一致。B鏈中含有高達25%的螺鏇區,它有凝集素的活性。B鏈兩條寡糖鏈末端(GicNac)2(Man)8和(GicNac)2(Man)7甘露糖殘基可以與網狀內皮細胞,特別是巨噬細胞結合,後者細胞表面富含甘露糖受體,可以優先攝取蓖麻毒素,這對於毒素髮揮生物功能有重要作用。
分離純化
蓖麻毒蛋白已被廣泛用於“導向藥物”的製備(即免疫毒素),將蓖麻毒蛋白套用於生物農藥方向也已受到廣泛關注與研究,因此,蓖麻毒蛋白的提取純化具有重要意義,研究提高蓖麻毒蛋白提取率的方法,並用於大規模生產之中,是許多專家正在進行的一項重要研究。從蓖麻種子或蓖麻餅粕中提取蓖麻毒蛋白粗提液的方法主要有:乙酸溶液提取法;磷酸緩衝溶液提取法;硫酸銨沉澱法和透析法等。隨著基因技術的發展,利用基因克隆方法獲得Ricin亦有成功報導。
(1)乙酸提取法,稱取適量蓖麻餅在研缽中研碎(研碎置於冰浴中,防止蓖麻毒蛋白在研碎過程中變性),加入適量乙醚以脫去脂肪。浸泡20min後,用四層紗布過濾,以濾去含脂乙醚。將脫脂後的蓖麻餅在室溫下晾乾,用冰醋酸在4℃下分別浸泡20h,用紗布過濾得粗提液。
(2)酸緩衝液提取:用磷酸緩衝液浸泡蓖麻餅處理樣品20h後,用聚乙二醇吸水,經5000r/min離心機離心15min,取上清液測定蓖麻毒蛋白含量。
(3)硫酸銨沉澱和透析法:在濾液中分別加入固體硫酸銨70%飽和度,5000r/min離心20min,所得深沉物先對蒸餾水透析,再對pH7.7的HCl-Tris緩衝液透析。透析後的樣品用相對分子質量為6000的聚乙二醇覆蓋1-2天,直至把樣品中的水分吸乾得粗蓖麻毒蛋白。收集所得粗蓖麻毒蛋白,用紫外分光光度計測定蓖麻毒蛋白含量。利用蓖麻毒素和凝集素對D-半乳糖基結合的特性,採用瓊脂糖Sepharose4B吸附,用半乳糖洗脫,再用纖維素離子交換柱層析或葡聚糖凝膠過濾可進一步純化得到蓖麻毒素。高寶岩將適量去殼蓖麻籽和石油醚加入到食品粉碎機中進行粉碎脫脂,過濾除去石油醚,用乙醚洗滌濾餅,所得脫脂蓖麻粉置於用醋酸調整pH=3.8的醋酸水溶液中浸取12h,所得上清液對蒸餾水透析24h,然後對pH=6.5磷酸緩衝溶液透析24h,透析液經25000r/min超速離心30min,上清液為蓖麻毒蛋白粗提液。然後將蓖麻毒蛋白進行柱層析分離純化,毒蛋白粗提液經瓊脂糖Sepharose4B分離得到純品。鄭成等通過乙酸提取、硫酸銨沉澱、透析,製備蓖麻粗蛋白,用紫外可見分光光度計測定蓖麻毒蛋白的含量,還使用高效液相色譜進行了分析,提取率較高,結果較滿意。
(4)隨著基因工程技術的發展,已能利用基因克隆的方法製備Ricin。從蓖麻籽中提取信使核糖核酸(MRNA),建立CDNA文庫,通過克隆獲得Ricin。由於基因轉導的高效性、較高的表達水平和完整的翻譯後加工過程,此系統適用於製備臨床治療用的Ricin。
檢測
蓖麻毒蛋白分析檢測尚缺乏簡單、快速、準確的定量分析方法,通用的方法如紅血細胞凝集法、280nm紫外吸收法,僅達目視比較半定量分析,還不適用於工業化規模的產品控制分析,更缺乏同時檢出能力。鄭成、高寶岩用高效液相色譜法在色譜柱150×4.6mm,5μm鍵合C4固定相,水、乙腈混合流動相,流速1mL/min,紫外檢測器測定波長280nm的色譜條件下,同時測定蓖麻毒蛋白和蓖麻鹼,效果令人滿意。試紙膜免疫層析法是一種快速免疫檢測技術。它是繼同位素、酶、螢光素等三大傳統標記技術以及各種免疫檢測法之後,套用金標記,帶色乳膠及免疫層析原理結合而發展起來的具有專一性強,靈敏度高,操作簡單,反應快速及經濟實用等特點。這種檢測法把以往對毒素的檢測法(包括需要數天的動物毒性測定及進行電泳等理化分析)以及廣泛採用的酶聯免疫吸附法(即ELISA法,一般測定時間4-8h),簡化到一步法,其實用性明顯。
毒理學
毒性
蓖麻毒蛋白是蓖麻毒素中毒性最強的一種,對各種哺乳動物都有毒。家畜中,兔和馬較敏感,羊和雞等較不敏感。兔(肌肉注射)半數致死劑量LD50為4.1μg/kg,小鼠(腹腔注射)LD50為10μg/kg,人經口致死量為0.15-0.2g,靜脈致死量為20mg。蓖麻毒蛋白是一種細胞毒素,對小白鼠有毒,但對斜紋夜峨無毒,可能是小鼠腸胃能吸收蓖麻毒素,而昆蟲的腸胃細胞不易於吸收大分子。作用機理
 蓖麻毒蛋白
蓖麻毒蛋白每個細胞可以結合106-108個蓖麻毒蛋白分子。Tonevitsky等用膜脂中含GMI脂質體作為細胞膜模型證明了蓖麻毒蛋白B與膜作用是通過兩個區域:一個半乳糖結合位點和一個疏水區,一個二亮氨酸基序或K-K/R-F/Y-F基序都可以充當內吞信號。一些生長因子、轉鐵蛋白、脂蛋白和激素都可能參與蓖麻毒蛋白分子的吸收。進入細胞後,大部分又回到細胞表面,僅有少量在高爾基體或溶酶體的作用下被裂解成兩條單鏈即A鏈和B鏈。Endo等研究表明它是以酶的方式直接作用於60S大亞單位28SrRNA,水解4324位點腺嘌呤N-糖苷鍵,使其喪失RNA酶的抗性而被水解,從而抑制蛋白質合成。一般生理狀況下1分子蓖麻毒蛋白A鏈能在1分鐘內脫去1000-2000個哺乳動物核糖體的腺嘌呤。Endo等研究表明,Ricin是以酶的方式直接作用於60S大亞基。A鏈具有RNAN-糖苷酶活性,特異性地水解真核細胞核糖體28SrRNA的第4324位腺苷酸的糖苷健,釋放出一個腺嘌呤鹼基,使核糖體失活。Tonevitsky等用膜脂中含GM1脂質體作為細胞膜模型,證明了RTB與膜作用是通過兩個區域:半乳糖結合位點和一個疏水區。B鏈與膜結合,幫助A鏈進入細胞。
Ricin通過RTB(B鏈)連線在細胞表面含有半乳糖末端的糖蛋白和脂蛋白上進入細胞,每個細胞可以結合106到108個Ricin分子。Ricin最可能通過包被小窩和小泡進入細胞。一些脂蛋白、轉鐵蛋白質、生長因子和激素都可能參與Ricin分子的吸收作用。Ricin分子只有通過高爾基體(TGN)進入胞質溶膠中才能發揮毒性作用。Ricin進入細胞後,部分被溶酶體降解,部分又回到細胞表面,僅少量進入到高爾基體中(對Ricin如何進入TGN還不了解)。通過高爾基體逆向轉運,Ricin到達內質網。一旦進入胞質溶膠,RTA(A鏈)便會催化核糖體的脫嘌呤作用,1min便可導致1500個核糖體失活,從而抑制蛋白質合成。Ricin不僅具有N—糖苷酶活性,使核糖體失活,當RTB與細胞膜表面的受體結合後,信號從膜受體到核的轉導過程中,還能誘導細胞凋亡、細胞因子的產生和脂質體過氧化等毒性作用。
尚沒有關於RTA鏈對其天然底物28S核糖體的作用模型,但人們已用底物類似物和人工合成寡核苷酸序列對其作用機制進行研究,認為底物本身的構象對酶的識別和催化很關鍵,脫嘌呤特異靶位點是一個富含嘌呤鹼基的,並具有12或14個核苷酸序列。處於中央的GAGA四核苷酸形成髮夾結構,這種四環結構能被A鏈所攻擊,其中第一個A(腺嘌呤核苷酸)便是要脫去的A(如圖2)。雖識別核糖體機制尚有待於研究,但人們可藉助於底物類似物FMP和APG的A鏈複合物晶體模型,提出一個合理的脫嘌呤機制。公認的RTA作用機制符合於RTA結構和動力學分析。底物結合於活性部位裂隙中,要脫去的A(腺苷)與Tyr80和Tyr123重疊形成”三明治”結構。
基因脫毒
生物技術的發展及基因沉默技術的出現,為蓖麻脫毒問題的解決提供了新的方法。基因沉默理論認為,導入與內源基因有較高同源性的基因可加強內源基因的沉默。AngelSM和HamiltonAJ等的研究也表明,轉入重複DNA片段引起內源基因近100%的轉錄後沉默。這為轉基因沉默內源基因提供了更加高效的方法。共抑制(co-suppression)指基因間相互作用引起的失活現象,當引入一個與植物內源基因有部分同源性的高效基因表達結構時,不但外源基因未能按預期方式進行高效表達,反而抑制了內源基因的表達。這些研究結果表明,重複抑制比單基因抑制效果好,反義基因比正義基因更能抑制同源基因的表達。臨床表現
中毒後數小時出現症狀。早期有精神不振,噁心嘔吐,腹痛、腹瀉、便血;繼則出現脫水、血壓下降,休克嗜睡;嚴重者可出現抽搐、昏迷,牙關緊閉;最後因循環衰竭而死亡。少數病人可出現發燒、黃疸、便血、蛋白尿、無尿或血尿,終因酸中毒、尿毒症而死亡。該毒素易損傷肝、腎等實質器官,發生出血、變性、壞死病變。並能凝集和溶解紅細胞,抑制麻痹心血管和呼吸中樞,是致死的主要原因之一。雖然在嚴重情況下許多器官可能會受到影響,蓖麻毒素中毒的主要症狀仍取決於接觸方式與中毒量。吸入蓖麻毒素中毒的最初症狀可能在接觸後8小時內才會出現。如果是吞咽蓖麻毒素中毒,最初症狀一般在不到6小時內出現。
吸入:在吸入大量蓖麻毒素後的幾小時內,可能的症狀是呼吸窘迫(呼吸困難)、發燒、咳嗽、噁心和胸悶。接著是大量出汗,且肺中積聚液體(肺水腫)。
這會使呼吸更加困難,且皮膚發紫。過多的肺中積聚液體要透過X射線或用聽診器聽胸部才能診斷。最後,將可能出現低血壓與呼吸衰竭,從而導致死亡。知道接觸了蓖麻毒素時,若吸入蓖麻毒素後12小時內開始出現呼吸症狀,應尋求醫。
吞咽:如果吞咽大量蓖麻毒素,將會出現帶血性嘔吐與腹瀉。結果會嚴重脫水,然後是低血壓。其他症狀可能包括出現幻覺、癲癇和尿中帶血。在幾天內,肝臟、脾與腎可能會停止工作,導致死亡。
皮膚與眼接觸:粉末狀或薄霧狀的蓖麻毒素可導致皮膚與眼睛變紅和疼痛。
救治措施
立即用高錳酸鉀或炭末混懸液洗胃,隨繼口服鹽類瀉藥及高位灌腸等急救措施,以排出未被吸收之毒物。口服乳汁、雞蛋清及阿拉伯膠,以保護胃黏膜。如出現昏迷、嗜睡等症狀時,可皮下注射可拉明、樟腦磺酸鈉等,必要時可用洋地黃製劑。如因大量嘔吐、嚴重腹瀉而失水時,應及時大量靜滴5%葡萄糖生理鹽水或低分子右鏇糖酐,以防止脫水導致休克,並有利於稀釋與排出毒素。注意糾正酸中毒,發生驚厥時給予苯巴比妥等鎮靜劑。有條件時,儘早注射抗蓖麻毒血清和輸血等治療措施。
套用
抗癌藥物
蓖麻毒蛋白(ricin)是一種植物毒蛋白,具有相當明顯的抗腫瘤作用,它們能通過抑制蛋白質合成來殺死癌細胞。蓖麻毒蛋白抗癌機理主要是:它能強烈地抑制各種癌細胞的蛋白合成,中度抑制DNA的合成,而對RNA的抑制則較弱。另一個重要的藥理作用是:它具有很強的抗原性,可經各種途徑進入機體,並可產生抗體和過敏反應,由於蓖麻毒蛋白能產生細胞毒作用,所以它能抑制巨噬細胞等參與免疫功能。但由於蓖麻毒蛋白的毒性極強,在使腫瘤減退的同時常伴隨有體重增加、水腫、血中蛋白質減少等毛細血管滲漏綜合症及神經性毒性反應,因此,限制了它在腫瘤化療中的套用。Ricin能標靶到特殊的細胞,如癌細胞。早在Ricin作用機制被了解之前就已經發現了其抗腫瘤特性。1951年,Mosinger報導Ricin對大鼠肉瘤的效用。1970年,Lin等報導Ricin可治療艾氏腹水瘤。對小鼠艾氏腹水癌的生長有預防作用。1980年,Fodstad和Pihl驗證Ricin和阿黴素聯合對接種白血病L1210細胞的小鼠的效果,他們發現了協同作用的效應及壽命延長198%,說明Ricin與阿黴素聯合套用的協同作用。研究證明:Ricin是一種新型的抗癌新藥,對人體頭頸部腫瘤細胞具有一定療效,對惡性黑色毒癌、結腸癌、宮頸癌、卵巢瘤有很好的療效,是一種新型很有希望的抗癌藥物。蓖麻籽中所提取的蓖麻毒素與胃癌抗體結合對腫瘤有抑制作用。
儘管在抗癌方面的套用受到普遍關注,但是蓖麻毒蛋白對細胞殺傷是非特異性的,在殺傷腫瘤細胞的同時常伴隨著體重增加、水腫、血中蛋白減少等毛細血管滲漏綜合症及神經毒性作用,而且蓖麻毒蛋白對免疫功能有強抑制性,因此需採用物理、化學、基因工程手段對蓖麻毒蛋白進行改性研究,以求提高其抗癌活性及靶向性並降低其副作用。Morris等對蓖麻毒蛋白A鏈做了缺失20個,5個或2個胺基酸的突變體,並測定了突變體的生物活性,為揭示蓖麻毒蛋白A鏈的結構功能關係提供了大量可靠的實驗數據。Lambert報導了通過使蓖麻毒蛋白與胎球蛋白分離的親和配體結合,並將產物進行親和層析,獲得B鏈被配體結合的A鏈活性未被阻斷的蓖麻毒蛋白。Pattrick等分別採用甲氧基聚乙二醇(MPEG)和聚乙二醇(PEG)修飾蓖麻毒蛋白,發現修飾物能在同等水平上抑制蛋白質的合成,細胞毒性卻降低了兩倍。ChristiansenSP用蓖麻毒蛋白-mAb35治療重症肌無力和斜視。Astashkin等經實驗證實無論是RTB還是整個毒蛋白分子都會增加Ca2+在人體淋巴細胞內的濃度,其可能與激活細胞膜磷酸肌醇的代謝和形成肌醇-1,4,5-三磷酸鹽有關,這也對RTA滲透進入細胞質至關重要。
Bushueve和Tonevitsky實驗測定出在2個不同的pH值下,7.5和5.0時蓖麻毒蛋白和其次級結構的穩定性顯著不同,當pH值從5.0升高時,蓖麻毒蛋白分子和RTA的穩定性在增加,而RTB的穩定性在下降。
蓖麻蛋白治療肝癌減輕了骨髓抑制的副作用,經肝動脈局部灌注5-氟脲嘧啶、絲裂黴素或順鉑等化療藥物,同時合併用明膠海綿、碘油等栓塞腫瘤供血動脈,或在碘油中加入少量化療藥物進行化療栓塞,均取得一定效果。但肝動脈灌注化療藥物一般量較大,這樣在殺滅癌細胞的同時,也損傷了正常的組織細胞,造成藥物性肝損害及骨髓抑制。蓖麻蛋白等植物毒素的特點就在於其用量極微就足以殺滅癌細胞,從而使上述困難有所改觀。另一方面,肝癌介入化療雖然比靜脈化療減少了消化道反應等副作用,但骨髓抑制現象仍然比較嚴重,350餘例肝癌病人中血小板、白細胞、血色素減少的發生率接近100%,有少數病人因白細胞數不能回升,而失去了再次介入治療的機會,也有部分病人原發性肝癌已基本消滅,但卻死於轉移,說明病人免疫功能受到抑制。通過實驗發現,蓖麻蛋白對裸鼠骨髓的抑制輕微,白細胞數、血紅蛋白量與對照組比較差別無顯著性,胸骨片上亦無白細胞抑制表現明顯優於絲裂黴素。
魯小青等探索出一種療效顯著的新型肝癌介入治療新藥,對蓖麻毒蛋白糖脂脂質體包封物進行了有關實驗研究:蓖麻毒蛋白與糖脂脂質體-半乳糖神經醯胺(galactcsylceramideliposomes,簡稱GCL)進行包封后形成的蓖麻毒蛋白糖脂脂質體包封物(R-GCL)作為設計藥物進行實驗研究。蓖麻毒蛋白作用後肝癌細胞的存活率46.6%。說明蓖麻毒蛋白糖脂脂質體包封物在體外實驗中對肝細胞有明顯的殺傷作用。龔承友等為了觀察蓖麻蛋白碘化乳劑介入治療肝癌的療效,用人肝癌裸小鼠皮下移植瘤為組織來源,植入實驗裸鼠肝內建立裸鼠肝癌模型,以瘤體內注入生理鹽水作對照,比較蓖麻蛋白(4μg/kg)碘油乳化蓖麻蛋白(8μg/kg)絲裂黴素(500μg/kg)(3種劑型用量均為半數致死量的三分之一)肝癌瘤體內注射後的療效,觀察腫瘤生長抑制率、甲胎、血像及骨髓改變。結果證明,蓖麻毒蛋白對肝癌的治療作用明顯,為蓖麻毒蛋白瘤內注射及乳化後肝動脈導向治療提供了實驗依據。
在1900年,德國藥物學家Ehrlicb就提出使藥物定嚮導向靶細胞而提高治療的專一性的構想,即用載體定向靶效應劑,如毒素、放射性核素或化療藥物等攜帶到病灶部位,在那裡發揮作用,而對正常細胞損傷很小。1975年,單克隆抗體(McAb)的問世為導向治療提供大量可選用的載體,使其向實用階段的目標前進了一大步。導向藥物包括抗體與毒素的連線,亦稱免疫毒素(IT,Immunotoxin)。免疫毒素有兩類:一類是抗體與完整毒素相連,另一類是抗體與毒素A鏈相連。蓖麻毒蛋白作為IT中最常用的毒素之一,伴隨著腫瘤導向治療的研究受到了普遍的重視。在美、法和中國進行的臨床試驗證實,蓖麻毒素對人體細胞癌的頭頸部腫瘤有一定療效,對惡性黑色素瘤、結腸癌、乳房癌、宮頸癌、胃癌等有較好療效。
大多數抗癌藥物作用於處於分裂期的細胞,因此對於分裂快的細胞也有殺傷作用,為此人們設計了將具有特異性導向性的單克隆抗體與高效的毒性彈頭的蛋白交聯為“導向藥物”,以改善抗癌藥物的臨床效果,組成的免疫導向毒素,既有單克隆體的識別功能,又有毒素的殺傷功能,並且能專一性殺傷靶瘤細胞,不損傷正常細胞,為蓖麻毒素用於惡性腫瘤的治療開闢了一條新途徑—腫瘤導向治療,被稱之為“生物飛彈”。免疫毒素的“彈頭”主要是各種生物來源的毒素。核糖體失活蛋白是常用的彈頭成分,其中蓖麻毒蛋白以其毒性強而被廣泛套用,其已被大量的套用於製備抗體導向的抗腫瘤藥物去定向的殺死腫瘤細胞,它們以酶催化方式發揮作用,毒性很強。蓖麻毒素具有N-糖苷酶活性,能特異地水解真核細胞28SrRNA第4324位上的腺嘌呤鹼,使其真核細胞60S核糖體失活,抑制蛋白質合成。因此,Ricin的研究已成為熱點。這是因為Ricin可構建免疫毒素,用以治療腫瘤。而且需求量與日俱增。
Ricin作為趨靶藥物在動物腫瘤模型及體外細胞培養中均能顯示其特異性的抗腫瘤能力,但臨床使用還存在很多問題,如對腫瘤的專一性和強殺傷能力二者不可兼得,因尚未找到真正的腫瘤特異性抗原,專一性不強。在血液循環中還可能被抗原所中和而導致失活等。
生物農藥
化學農藥、化肥等化學製品對植物產品和人類生存環境的污染問題是亟待解決的重大課題,也是實現中國農業可持續發展的主要障礙之一。利用易降解、對作物安全的植物源殺蟲劑代替有機殺蟲劑被很多植物保護專家們認為是解決這一問題的良好途徑。為此,開發和套用植物源農藥已成為各國所追逐的目標。生物殺蟲劑由於對人畜毒性很小和環境污染小得到廣泛的研究和開發。蓖麻毒蛋白作為生物農藥殺蟲後在蟲體內可以自然分解,不再危及下級食物鏈,可以解決農藥殘留的污染問題,因此蓖麻毒蛋白在生物農藥方面有著獨特的優勢和廣闊的套用前景。但是針對蓖麻毒蛋白的研究主要集中於醫學套用研究,特別是用於生物飛彈藥的開發研究,而針對生物農藥方面的研究還比較少。趙建興等用不同溶劑提取蓖麻抽提物對天幕毛蟲、桃蚜及小菜蛾進行殺蟲試驗,結果發現對天幕毛蟲幼蟲和桃蚜的殺滅活性物質是蓖麻鹼和蓖麻毒蛋白,毒蛋白主要是觸殺作用。尹秀玲利用蓖麻子和根葉的蓖麻毒素成分作為殺蟲劑,套用於農作物的殺蟲,取得了一定的效果。但由於蓖麻毒蛋白的分子量較大,對大多數昆蟲來說,難以在RTB的幫助下使RTA進入昆蟲細胞而發揮其毒性,有些利用蓖麻提取物殺滅昆蟲的研究,其主要殺滅成分為蓖麻鹼而非蓖麻毒蛋白,因此在保留蓖麻毒蛋白生物活性的前提下將其進行化學修飾,包括酶法修飾,降低其分子量,並篩選合適的導入試劑,可望發揮蓖麻毒蛋白的殺蟲效力。
中國已直接用蓖麻籽和它的根、莖、葉製成植物殺蟲劑,套用於實踐中取得了很好的效果。蓖麻毒素也可套用到滅鼠藥的研發上。蓖麻毒素中的蓖麻毒蛋白能阻擋或抑制老鼠體內蛋白質的合成,一個Ricin分子進入老鼠細胞內,就足以使其整個細胞的蛋白質合成停止而死亡,蓖麻變應原是一種強烈的過敏性物質,與Ricin及蓖麻鹼具有協同效應。老鼠食用由蓖麻毒素製成的鼠藥後,即可因細胞中毒,呼吸窘迫及組織器官功能衰竭致死。
Ricin作為植物源農藥的開發利用可分為兩方面:一是直接利用,即對蓖麻毒素進行粗提取後,直接加工成可利用的製劑。這種利用方式的主要優點是能夠發揮粗提物中各種成分的協同作用,而且投資少,開發周期短。二是間接利用,即研究Ricin的結構、作用機制、結構與活性間的關係,再進行化學修飾或人工模擬合成篩選,從中開發新型植物源農藥製劑。間接利用是除中國以外的其他國家對Ricin作為植物源農藥研究開發的重點,也是中國植物源農藥研究發展的方向。
其它
蓖麻毒蛋白分子能專一的與含有半乳糖末端殘基的多糖或糖蛋白結合,根據親和吸附的原理,蓖麻毒蛋白可用於分離純化相應糖基的糖蛋白、糖脂和多糖;同時還可以用來研究細胞表面糖的分布,從而成為研究細胞膜的有效方法。此外,蓖麻毒蛋白是一種核糖體失活蛋白,它也是理論上研究核糖體結構和功能的重要工具。另外,李淑華用提取的蓖麻蛋白配製膠粘劑,利用蛋白質高分子溶液遷移至木材表面,使蛋白質高分子極性基團接近木材表面的極性基團,當二者官能團之間距離與分子間力作用半徑大小相當時,產生結合力。蓖麻毒蛋mAb35是一種免疫毒劑,由蓖麻毒蛋白和作用於橫紋肌煙酸型乙醯膽鹼受體的單克隆抗體結合而成。局部套用直接作用於橫紋肌,導致長期的肌力減弱,因此可用於治療各種肌張力異常。蓖麻毒蛋mAb35主要作用於成熟的肌纖維,不影響成肌細胞的功能,因此導致的肌肉損害可以再生。眼外肌也是一種橫紋肌,蓖麻毒蛋白mAb35作用於特定的眼外肌,可引起長期的、劑量依賴性的張力改變,從而矯正斜視患者的眼位。
