
分類寄生
| 復殖目 Digenea | Digenea後睪科 Opisthorchiidae | 支睪屬 Clonorchis | 華支睪吸蟲 C.sinensis | 肝膽管 |
| | 異形科 Heterophyidae | 異形屬 Heterophyes | 異形異形吸蟲 H.heterophyes | 腸管 |
| | 片形科 Fasciolidae | 薑片屬 Fasciolopsis 片形屬 Fasciola | 布氏薑片吸蟲 F.buski 肝片吸蟲 P.hepatica | 小腸 肝膽管 |
| | 並殖科 Paragonimidae | 並殖屬 Paragonimus 狸殖屬 Pagumogonimus | 衛氏並殖吸蟲 P.westermani 期氏狸殖吸蟲 P.skrjabini | 肺(或腦) 皮下(或肝) |
| | 裂體科 Schistosomatidae | 裂體屬 Schistosoma | 日本裂體吸蟲 S.japonioum | 門脈系統 |
| | 棘口科 Echinostomatidae | 棘隙屬 Echinochasmus | 日本棘隙吸蟲 E.japonicus | 小腸 |
吸蟲
吸蟲(trematoda)
亦作fluke。
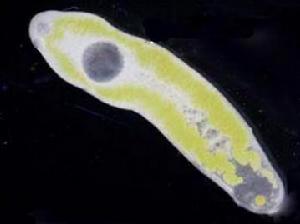 吸蟲
吸蟲扁形動物門(Platyhelminthes)吸蟲綱(Trematoda)動物,近6,000種,世界性分布。體長5公厘(0.2吋)至10公分(4吋)。寄生於各類脊椎動物,常寄生於魚、蛙和龜體;但也寄生於人體和無脊椎動物(如貝類、甲殼類)。外寄生、內寄生或半外寄生(吸附於口腔壁、鰓或泄殖腔),有的需兩個或兩個以上宿主。兩側對稱。體復以非細胞的角質層。多數扁平,葉狀或帶狀,但有的橫切面呈圓形。腹面有吸盤及鉤、刺用以吸附。體軟,所有器官周圍充以海綿狀結締組織(間充質);有或無循環系統。消化系統為一簡單的囊,口在前端或腹面中央。通常無肛門,但有的具一或兩個肛孔。神經系統有一對前神經節和3對縱行的神經索。多數雌雄同體。卵生或產出幼體、成體。
血吸蟲寄生於多數脊椎動物,寄生人體的有3種︰埃及裂體吸蟲(Schistosoma haematobium, 即埃及血吸蟲)生活在膀胱靜脈內,主要分布於非洲、南歐和中東。卵穿過靜脈壁進入膀胱,隨尿排出。幼蟲在中間宿主螺類(主要為Bulinus屬和Physopsis屬)體內發育。成熟幼蟲通過皮膚或口進入終宿主人體內。曼森氏裂體吸蟲(S. mansoni, 即曼氏血吸蟲)在大、小腸靜脈中,主要分布於非洲和南美洲北部。卵隨糞便排出。幼蟲進入螺體,再通過皮膚回到終宿主人體內。日本裂體吸蟲(S. japonicum, 即日本血吸蟲)主要見於中國大陸、日本、台灣、東印度群島和菲律賓,除人外,還侵襲其他脊椎動物,如家畜和大鼠等。中間宿主是釘螺屬(Oncomelania)軟體動物。成蟲在腸系膜靜脈中,有些卵隨血流進入各器官,引起各種症狀,如肝腫大。嚴重時造成宿主死亡。在非洲和東亞有數百萬人得血吸蟲病。肝片吸蟲(Fasciola hepatica)致綿羊和其他家畜的肝吸蟲病。人食用未經烹煮的水田芹等而後可被侵染。華枝睪吸蟲(Opisthorchis sinensis或Clonorchis sinensis)寄生在人、貓、狗等多種哺乳動物的肝臟膽管內,第一中間宿主是螺,第二中間宿主是淡水魚。貓後睪吸蟲(Opisthorchis felineus)的終宿主包括人、貓和狗等,也需要魚作為第二中間宿主。
吸蟲(trematoda)屬扁形動物門的吸蟲綱(Class Trematoda)。在人體中寄生的吸蟲均隸屬於復殖目(Order Digenea),稱為復殖吸蟲(digenetic trematode),其基本結構及發育過程略同。
大多數復殖吸蟲成蟲外觀呈葉狀、長舌狀。背腹扁平,兩側對
稱;通常具口吸盤(oral sucker)及腹吸盤(acetabulum),內部結構如下:
體壁組織 吸蟲成蟲體表有皺褶、凸起、陷窩、體棘、感覺乳突等,其形態、數量、分布等因不同蟲種、不同部位而異。體壁由體被(tegument)與肌肉層構成。中間為實質組織(parenchymal tissue),消化、生殖、排泄、神經系統等分布於其中,缺體腔。體被曾被認為是無活力的,只是保護作用的“角皮 ”(cuticle)。實際上,它是具有代謝活力的合胞體(syncytium)。從外到內由外質膜(external plasma membrane)、遠端胞漿(distal cytoplasm)區、基質膜(basal plasmamembrane)組成。與外質膜聯合在一起的是表面外膜(furface coat),也稱糖萼(glycocalyx),它是糖蛋白與糖脂上的糖殘支。遠端胞漿區內布滿基質,感覺器位於其中,它的纖毛伸出體表,另一端有神經突(nerve process)與神經系統相通。在遠端胞漿區中還有線粒體(mitochondrion)及分泌顆粒(secretory granule)體棘位於基質膜之上,基質膜之下為基層(basement layer)及肌肉層。肌肉層由外環肌(circular muscle)與內縱肌(longitudinal muscle)組成。肌肉層之下為近端胞漿區(proximal cytoplasm),也即細胞體區(cyton region),內有核、內質網(endoplasmic reticulum)、高爾基複合體(Golgi complexes)、線粒體、分泌顆粒、各種小泡(vesicles)。修復和維持遠端區的物質都在此合成並通過窄小的胞漿連結部(cytoplasmic connective)輸送到遠端胞質區。各種蟲及不同發育階段的體被不盡相同,但總的來說其功能是保護蟲體、吸收營養及感覺。
消化系統 復殖吸蟲有不完全的消化道,包括:由肌性口吸盤圍繞的口、前咽(prepharynx)、咽(pharynx)、食管(esophagus)和腸管,後者常分為兩個腸支(cecum)。口、咽、食管構成前腸(foregut)。前腸最裡層類似體被結構,即其合胞體細胞由近端區及遠端區構成。腸支內壁為單層細胞層,其胞質伸出具漿膜的絨毛樣褶以擴大吸收面積。前腸及腸支雖具有吸收及消化動能,但主要是在前腸及腸管前部進行。吸蟲的消化是一種典型細胞外消化,即細胞分泌酶將食物消化,然後由細胞吸收。
生殖系統 復殖吸蟲除血吸蟲外都是雌雄同體(hermaphrodite),即同一蟲體內具雌、雄兩套生殖系統,通常雄性生殖系統早熟於雌性生殖系統,這似乎能減少自我受精機會。雄性生殖系統包括睪丸(testis)、輸出管(vas efferens)、輸精管(vas deferens)、儲精囊(seminal vesicle)、前列腺(prostatic gland)、射精管(ejaculatory duct)或陰莖(cirrus)、陰莖袋(cirrus pouch)等組成。在某些蟲種,一些結構,如前列腺、陰莖袋、陰莖等可能會缺失。雌性生殖系統由卵巢(ovary)、輸卵管(oviduct)、卵模(ootype)、梅氏腺(Mehlis gland)、受精囊(seminal receptacle)、勞氏管(Laurer’s canal)、卵黃腺(vitellaria)、卵黃管(vitelline duct)、總卵黃管(commom vitelline duct )、卵黃囊(vitelline reservior)、子宮(uterus)、子宮末段(metraterm)等組成。雌雄生殖系統的遠端均開口於生殖竇(genital atrium)。交配時,陰莖插入子宮末段,精子游進受精囊並儲存於此,卵的受精一般在輸卵管進行,卵黃細胞顆粒在梅氏腺分泌物的作用下,卵殼前體鞣化(tan)為較硬卵殼,或聯粘成二硫化物,使之形成更有彈性卵殼。之後卵進入子宮經生殖孔排出。復殖吸蟲的生殖系統最為發達,每日產卵量多,所需營養物質也最多,合成代謝與能量代謝也最旺盛。各種進入蟲體物質都在生殖系統進行代謝與消耗,同樣地,殺蟲藥也會在此積聚造成結構與功能損傷,甚至引起死亡。
排泄系統 吸蟲排泄系統由焰細胞(flame cell)、毛細管(capillary tubule)、集合管(collecting tubule)與排泄囊(excretory bladder)組成,經排泄孔通體外。焰細胞與毛細管構成原腎(protonephron)單位。焰細胞的數目與排列可用焰細胞式(flame cell pattern)表示。它是吸蟲分類的重要證據。焰細胞有細胞核、線粒體、內質網等。胞漿內有一束纖毛,每一纖毛有兩根中央纖絲(fibril)與9根外周纖絲組成。活體顯微鏡觀察時,纖毛顫動象跳動的火焰,因而得名。纖毛顫動使液體流動並形成較高的過濾壓,促使含有氨、尿素、尿酸等廢物的排泄物排除體外。
神經系統 咽的兩側各有一神經節(ganglion),有背索相連(esophageal commissure)。每個神經節分別向前、後各發出背、腹、側3條神經乾(nerve trunk)分布於蟲體的背面、腹面、側面。向後的神經乾間在不同水平有橫索(transverse commissure)相連,使整個神經系統形成“梯子形”(ladder)。由神經乾發出的神經末梢到達口吸盤、咽、腹吸盤、生殖系統等器官及體壁外層感覺器。神經系統有乙醯膽鹼脂酶與丁醯膽鹼脂酶的活動,神經節中有神經分泌細胞的存在,說明神經系統機能相當活躍。
吸蟲的生活史
1.復殖吸蟲
復殖吸蟲的生活史都需經歷世代交替即有性世代(有性生殖,sexual generation)與無性世代(無性生殖,asexual generation)的交替。無性世代一般在中間宿主,如軟體動物腹足類(gastropod)的螺螄與斧足類的蚌中。有性世代大多在脊椎動物(終宿主)。復殖吸蟲生活史雖較複雜,但基本生活史類型包括卵(ovum)、毛蚴(miracidium)、胞蚴(sporocyst)、雷蚴(redia)、尾蚴(cercaria)、囊蚴(encysted metacercaria)、後尾蚴(從囊中脫出的幼蟲,metacercaria)與成蟲(adult)。卵從成蟲所寄生的器官排進宿主腔道並隨排泄物排出體外,在水中孵出毛蚴或被宿主吞食後才孵出毛蚴。毛蚴侵入螺螄淋巴系統或其它器官發育為胞蚴。胞蚴體內胚團分裂發育成多個雷蚴。雷蚴中的胚團再分裂發育為多個子雷蚴。在形態上雷蚴與胞蚴不同,雷蚴前端已具口、肌性的咽及短的腸支。胞蚴與雷蚴均靠體表吸取其周圍組織中的營養。雷蚴與子雷蚴中的胚團分裂發育為尾蚴。在不利環境,如寒冷季節,有些吸蟲的雷蚴不產生尾蚴而只產生雷蚴,並可連續數代。在血吸蟲也可見類似情況:子胞蚴不產生尾蚴而產生胞蚴。這種變換生殖現象稱為多胚繁殖(polyembryonic proliferation)。有些吸蟲缺雷蚴期或囊蚴期,而另一些吸蟲卻具有兩代以上的雷蚴期。吸蟲的感染期是尾蚴或囊蚴。囊蚴被宿主吞食後,後尾蚴脫囊而出,部分吸蟲後尾蚴及以尾蚴為感染期的吸蟲需移行才能到達適宜發育的定居部位。不同器官組織為蟲體提供不同發育期所需的營養物質,蟲體能識別不斷改變的連續刺激,使大部分蟲體能按一定移行途逕到達定居部位。不適宜宿主不能提供必需的營養物質及生理信號,因而出現異常的個體發生移行(ontogenetic migration),導致異常寄生或發育遲緩甚至死亡。
2.單殖吸蟲
單殖吸一般是直接發育,也就是從母體可以直接產生幼體。如三代蟲,幼體脫離母體後能在水中自由游泳尋覓寄主進行寄生。
吸蟲發育中基因的調控
復殖吸蟲生活史所有階段的基因是相同的,但在不同階段其基因表達是不同的,例如,負責雷蚴發育的基因在胞蚴發育階段是被抑制的,反之也然。其激活與抑制機制至今未明。
危害
 肝片吸蟲
肝片吸蟲吸蟲絕大多數是各類脊椎動物的寄生蟲病的病原,軟體動物等因被吸蟲的幼蟲期所寄生亦受損害。因此,人及各類經濟動物均可受到不同程度的危害。
單殖亞綱分單後盤目(Monopisthocotylea)及多後盤目(Polyopisthocotylea)。種類在千種以上,絕大多數是魚類的體外寄生蟲。它們通常以其後附著器的幾丁質結構插入被寄生部位的組織,破壞鰓及皮膚的組織,造成炎症,引起病變;及吸吮魚血、粘液,引起繼發性魚病。有時可導致魚苗的大批死亡。單殖吸蟲對中國淡水養殖魚類的危害,主要有三代蟲和指環蟲二屬種類所引起的鰓病和皮膚病。
復殖亞綱種類繁多,約有140餘科1400多屬、萬種以上,占吸蟲綱的大部分。其中一些種類是人體和經濟動物(珍貴動物、家畜、家禽、魚類及經濟貝類等)的吸蟲病病原。如腹口吸蟲目及前口吸蟲目的孔腸科等吸蟲是魚類寄生蟲,幼蟲期寄生在雙殼綱軟體動物如貽貝、珍珠貝、縊蟶等上,產生嚴重危害。在前口目中的裂體科、並殖科、雙腔科、片形科、同盤科、後睪科及棘口科等均有人體和家畜、家禽及魚類等經濟動物重要吸蟲病的病原。多種吸蟲病原可使家畜消瘦,甚至大批死亡。應根據各吸蟲生活史各階段的生物學特點,中間宿主(傳播媒介)的生活習性、吸蟲病原存在、散布的生態學及流行病學特點而採取綜合防治措施。
