
過程
間期
間期又分為三期、即DNA合成前期(G1期)、DNA合成期(S期)與DNA合成後期(G2期)。
1.G1期(first gap) 從有絲分裂到DNA複製前的一段時期,又稱合成前期,此期主要合成RNA和核糖體。該期特點是物質代謝活躍,迅速合成RNA和蛋白質,細胞體積顯著增大。這一期的主要意義在於為下階段S期的DNA複製作好物質和能量的準備。
2.S期(synthesis) 即DNA合成期,在此期,除了合成DNA外,同時還要合成組蛋白。DNA複製所需要的酶都在這一時期合成。
3.G2期(second gap)期為DNA合成後期,是有絲分裂的準備期。在這一時期,DNA合成終止,大量合成RNA及蛋白質,包括微管蛋白和促成熟因子等。
分裂期
M期:細胞分裂期。
細胞的有絲分裂(mitosis)需經前、中、後,末期,是一個連續變化過程,由一個母細胞分裂成為兩個子細胞。一般需1~2小時。
1. 前期(prophase)染色質絲高度螺鏇化,逐漸形成染色體(chromosome)。染色體短而粗,強嗜鹼性。兩個中心體向相反方向移動,在細胞中形成兩極;而後以中心粒隨體為起始點開始合成微管,形成紡錘體。隨著核仁相隨染色質的螺鏇化,核仁逐漸消失。核被膜開始瓦解為離散的囊泡狀內質網。
2. 中期(metaphase)細胞變為球形,核仁與核被膜已完全消失。染色體均移到細胞的赤道平面,從紡錘體兩極發出的微管附著於每一個染色體的著絲點上。從中期細胞可分離得到完整的染色體群,共46個,其中44個為常染色體,2個為性染色體。男性的染色體組型為44+XY,女性為44+XX。分離的染色體呈短粗棒狀或髮夾狀,均由兩個染色單體借狹窄的著絲點連線構成。
3.後期(anaphase)由於紡錘體微管的活動,著絲點縱裂,每一染色體的兩個染色單體分開,並向相反方向移動,接近各自的中心體,染色單體遂分為兩組。與此同時,細胞被拉長,並由於赤道部細胞膜下方環行微絲束的活動,該部縮窄,細胞遂呈啞 鈴形。
4.末期(telophase)染色單體逐漸解螺鏇,重新出現染色質絲與核仁;內質網囊泡組合為核被膜;組胞赤道部縮窄加深,最後完全分裂為兩個2倍體的子細胞。
以有絲分裂方式增殖的細胞從一次分裂結束到下一次分裂結束所經歷的過程。這一過程周而復始。細胞周期是50年代細胞學上重大發現之一。在這之前認為有絲分裂期是細胞增殖周期中的主要階段,而把處於分裂間期的細胞視為細胞的靜止階段 。1951 年霍華德等用P-磷酸鹽標記了蠶豆根尖細胞,通過放射自顯影研究根尖細胞DNA合成的時間間隔,觀察到P之摻入不是在有絲分裂期,而是在有絲分裂前的間期中的一段時間內。發現間期內有一個DNA合成期(S期),P只在這時才摻入到DNA;S期和分裂期(M期)之間有一個間隙無P摻入,稱為G2期,在M期和S期之間有另一個間隙稱為G1期,G1期也不能合成DNA。
細胞生命活動大部分時間是在間期度過的,如大鼠角膜上皮細胞的細胞周期內,間期占14000分鐘。分裂期僅占70分鐘。細胞周期各階段都有複雜的生化變化。間期是細胞合成DNA、RNA、蛋白質和各種酶的時期,是為細胞分裂準備物質基礎的主要階段。
在一個增殖的細胞群中,所有細胞並非是同步增殖的,它們在細胞周期運行中,可能有四種命運:
①細胞經M期又開始第二次周期;
②停止於G2期,稱為G2期細胞(R2),它受某種刺激後可進入周期;
③停止在G1期,稱為休止細胞或G期細胞,這類細胞受某種刺激後仍能進入周期,繼續進行有絲分裂;
④喪失生命力近於死亡的細胞,稱為丟失細胞,或稱不再分裂的細胞。繼續分裂的細胞沿著細胞周期從一個有絲分裂期到下一個分裂期。不再分裂的細胞離開了細胞周期不再分裂,最終死亡。
G1期
細胞體積逐漸增大,製造RNA(包括tRNA,mRNA,rRNA以及核糖體等)。RNA的合成又導致結構蛋白和酶蛋白的形成,這些酶又控制著形成新細胞成分的代謝活動。G1又分為G1早期和G1晚期兩個階段;細胞在G1早期中合成各種在G1期內所特有的RNA和蛋白質,而在G1晚期至S期則轉為合成DNA複製所需要的若干前體物和酶分子,包括胸腺嘧啶激酶、胸腺嘧啶核苷酸激酶、脫氧胸腺嘧啶核苷酸合成酶等,特別是DNA聚合酶急劇增高。這些酶活性的增高對於充分利用核酸底物在S期合成DNA是不可少的條件。
 細胞周期
細胞周期G1期持續時間變異很大,多數細胞的G1期較長,是與細胞需要增加質量有關。但在某些單細胞生物如大變形蟲、四膜蟲和多細胞生物的某些細胞(如海膽胚胎,小鼠胚胎細胞)則無G1期,中國倉鼠卵巢細胞的變異株無G1和G2期,以致M期和S期連線在一起。G1期的長短之所以變化很大,與G1期記憶體在一個校正點或阻止點(簡稱R點)有關。R點主要控制 G1期時間的長短。通過了此點,細胞就能以正常速度不受外界條件的影響而完成細胞周期的其他時期。因此,有人認為細胞的生長是在G1期R點上停止的,例如當細胞內環腺苷酸(cAMP)水平增高,細胞密度增加時,可阻止細胞從G1期向S期過渡,用嘌呤黴素抑制蛋白質合成或用放射線菌素D抑制RNA合成,也能延緩細胞從G1期進入S期。有人發現 G1期內能合成一種有觸發作用的蛋白質;它是不穩定的,極易被分解,故稱為v蛋白。v蛋白在G1細胞中達到一定水平時,細胞便可通過R點進入S期。
細胞進入G1期後,並不是毫無例外地都進入下一期繼續增殖,在此時可能會出現三種不同前景的細胞:①增殖細胞:這種細胞能及時從G1期進入S期,並保持旺盛的分裂能力。例如消化道上皮細胞及骨髓細胞等;②暫不增殖細胞或休止細胞:這類細胞進入G1期後不立即轉入S期,在需要時,如損傷、手術等,才進入S期繼續增殖。例如肝細胞及腎小管上皮細胞等;③不增殖細胞:此種細胞進入G1期後,失去分裂能力,終身處於G1期,最後通過分化、衰老直至死亡。例如高度分化的神經細胞、肌細胞及成熟的紅細胞等。
G0期
細胞周期的調節主要是通過G1期的阻留而實現的,G0期即指細胞處於阻留的狀態。細胞通過M期一分為二,有的可繼續分裂進行周期循環,有的轉入G0期。G0期是脫離細胞周期暫時停止分裂的一個階段。但在一定適宜刺激下,又可進入周期(圖1),合成DNA與分裂。G0期的特點為:①在未受刺激的G0細胞,DNA合成與細胞分裂的潛力仍然存在;②當G0細胞受到刺激而增殖時,又能合成DNA和進行細胞分裂。
S期
在這一階段完成DNA的合成以及合成與DNA組裝構成染色質等有關的組蛋白。DNA含量在此時期增加一倍。S期終結時,每一染色體複製成兩個染色單體(Hole,1979)。生成的兩個子代DNA分子與原來DNA分子的結構完全相同。一個人體細胞核直徑10~20微米,其中DNA含量為10克,如拉成一根DNA鏈,長度可達3米。哺乳類動物細胞S期一般為6~8小時。DNA的複製能在幾小時內完成,主要是由於DNA鏈分成許多的複製單位(複製子)(可多達10000個左右),它們可在S期的不同時間分別複製。另外,在S期內還有組蛋白的合成──組蛋白基因在G1-S期之間活化,組蛋白mRNA的轉錄增大,並在整個S期內連續進行。已合成的組蛋白使新合成的DNA很快轉為核組蛋白複合體。
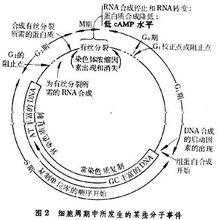
S期細胞含有一種因素能誘導DNA合成,用細胞融合實驗證明,G1細胞在與S期細胞融合後能加速其核內DNA複製的起點啟動。S期不同階段複製的DNA鹼基組成是不同的,早期複製的DNA富有G-C鹼基,晚期複製的DNA富有A-T鹼基,即常染色質比異染色質複製較早(圖2)。
G2期
是DNA複製結束和開始有絲分裂之間的間隙,在這期間細胞合成某些蛋白質和RNA分子,為進入有絲分裂提供物質條件。用放射標記的RNA前體和蛋白質前體示蹤,表明G2期進行著強烈的RNA和蛋白質的合成。假如破壞這些合成過程,細胞就不能過渡到M期。G2期合成的是染色體濃縮以及形成有絲分裂器所需的成分。有人認為G2期繼續完成從S期就開始的微管蛋白的合成,為M期紡錘絲的組裝提供原料。在G2晚期開始合成有絲分裂因子。在某些缺少G1期細胞中,G2期更為複雜,還要擔負起其他細胞G1期中所要完成的事件。也有少數情況,S期結束後立即開始有絲分裂,而不存在G2期。
M期
有絲分裂時期,是細胞形態結構發生急速變化的時期,包括一系列核的變化、染色質的濃縮、紡錘體的出現,以及染色體精確均等地分配到兩個子細胞中的過程,使分裂後的細胞保持遺傳上的一致性。M期分為前期、中期、後期和末期(見有絲分裂)。M期雖是形態變化最為顯著的時期,但其呼吸作用反而降低,蛋白質合成明顯下降,RNA合成及其他代謝周轉停止,這是由於有絲分裂期所需要的能量和其他基本物質均在間期內合成和貯備好了有關。
細胞周期中,細胞形態也發生一系列變化,從光學顯微鏡下可看到G1期細胞最小,細胞扁平而光滑,隨著向S→G2→M期的發展細胞逐漸增大,從扁平變成球形。掃描電鏡下可明顯看到各時期內細胞表面形態的變化,如微絨毛逐漸增加,這些變化和細胞內各種生化的和生理的周期性變化是有關的。
調控細胞周期中的許多生化事件是按一定順序,有條不紊地進行著,這和基因按一定順序表達密切相關。
影響因素
細胞周期內有兩個階段最為重要:G1到S和G2到M;這兩個階段正處在複雜活躍的分子水平變化的時期,容易受環境條件的影響,如果能夠人為的進行調控,將對深入了解生物的生長發育和控制腫瘤生長等有重要意義。
已發現很多體內因素可以激發或抑制細胞的增殖,例如多種激素、血清因子、多胺、蛋白水解酶、神經氨酶、cAMP、cGMP以及甘油二脂(DG)、 三磷酸肌醇(IP3)和Ca信使系統等等。細胞內cAMP濃度增加對細胞增殖有抑制作用,凡能使細胞內cAMP增高的因素都能抑制細胞的增殖,降低細胞生長速度;反之,凡能使細胞內cAMP含量下降的因素都能促進DNA的合成與細胞的增殖。細胞周期的各期中的cAMP含量也不相同(見表)。在中國倉鼠卵巢細胞株中,M期cAMP含量最低,M期後cAMP的水平增高三倍,從G1早期至G1晚期,cAMP水平降低到中等水平,直至S期仍維持低的水平(圖3)。
還有許多實驗指出cGMP 也對細胞增殖起調控作用,如將cGMP或雙丁醯cGMP加到休止在G1期的 3T3 細胞時,能誘導DNA含量的增加, 促進細胞的分裂。如提高細胞cGMP水平,就可促進細胞的有絲分裂,反過來,促進有絲分裂的藥物也能增加cGMP的濃度。
cAMP能抑制細胞的分裂,促進細胞的分化,cGMP則能抑制細胞分化,促進細胞增殖,在正常生長的細胞中,cAMP和cGMP維持在適當的水平,調節控制細胞周期的運轉。
抑素是細胞產生的一種小分子蛋白質或多肽,有的還含有糖或RNA。它無種屬特異性,但有細胞特異性,對同類細胞增殖有抑制作用並且可逆。當抑素含量達到一定濃度時可抑制同類細胞的增殖,抑素濃度下降則細胞增殖活躍。有人認為抑素作用的機制,在於它能激活細胞膜上的腺苷環化酶活性,提高細胞內cAMP的濃度,因而抑制細胞的增殖,也可能通過cAMP-依賴性蛋白激酶對蛋白質的磷酸化作用來影響調節基因的活動。
細胞周期也受機體調節系統的影響,例如肝再生就是由調節系統的作用加速肝細胞增殖。但是腫瘤細胞,由於宿主失去對它的調控,因而惡性增殖。在腫瘤治療中可套用細胞周期的原理,如G0期細胞對化療不敏感,往往成為日後癌症復發的根源,因而可通過調控機理的研究,誘發G0期癌細胞進入細胞周期,再合理用抗癌藥物加以殺滅,是防止癌轉移和擴散的重要調控措施,是細胞動力學中有理論意義和實踐意義的研究問題。
總之,至今所了解的細胞增殖調控的分子基礎還少,尚待進一步探索。
細胞分類
在體內根據細胞的分裂能力可把它們分為三類:①周期性細胞,如造血幹細胞,表皮與胃腸黏膜上皮的幹細胞。這類細胞始終保持活躍的分裂能力,連續進入細胞周期循環;②終端分化細胞,如哺乳動物成熟的紅細胞、神經細胞等高度分化的細胞,它們喪失了分裂能力,又稱終末細胞(end cell);③暫不增殖細胞群(G0期細胞),如肝細胞、腎小管上皮細胞、心肌細胞、甲狀腺濾泡上皮細胞。它們是分化的,並執行特定功能的細胞,在通常情況下處於G0期,故又稱G0期細胞。在某種刺激下,這些細胞重新進入細胞周期。如肝部分切除術後,剩餘的肝細胞迅速分裂。
DNA增殖特點
細菌DNA複製、RNA轉錄和蛋白質合成同時進行,這是細菌對快速生長的原因。
DNA複製不受細胞周期的限制,快速生長的細菌,在上一次細胞分裂結束時,細胞內的DNA經複製到一半進程,以保證迅速進行下一次分裂。
1、細胞周期是第一次分裂開始到第二次分裂開始所經歷的全過程。《普通生物學》陳閱增
2、細胞周期是第一次分裂結束到第二次分裂結束所經歷的全過程。《細胞生物學》瞿中和,王喜忠,丁明孝
兩者區別是細胞分裂起點的不同。現今普遍接受的是第二種說法。
口訣
以 植物細胞有絲分裂為例:
有絲分裂分五段
間前中後末相連
間期首先作準備
間期染體複製在其間
前期 兩消兩現一散亂
中期 著絲點聚赤道板
後期 絲牽染體兩極走
末期 兩消兩現 壁重建
註:動物細胞末期不生成細胞壁。
