
燈刷染色體
正文
形如燈刷狀,是一類處於伸展狀態具有正在轉錄的環狀突起的巨大染色體。常見於進行減數分裂的細胞中。因此它常是同源染色體配對形成的含有 4條染色單體的二價體。卵母細胞發育中所需的全部mRNA和其他物質都是從燈刷染色體轉錄下來合成的。燈刷染色體首次由W.弗勒明在1882年報導,但未肯定這是一種染色體。1892年J.呂克特對在鯊魚卵母細胞中的這種巨大染色體的結構進行了研究,並因其形狀酷似歐洲當時擦洗煤油燈罩的燈刷,取名為燈刷染色體。50年代在H.G.卡倫等人改進了研究技術後,燈刷染色體的研究才蓬勃發展起來。
燈刷染色體在生物界分布廣泛,但出現的時期不同。大多數動物的燈刷染色體出現在卵母細胞第一次減數分裂的雙線期,例如軟體動物、節肢動物中的甲殼類和大多數昆蟲類、毛顎動物、棘皮動物和脊索動物。有些動物燈刷染色體出現在精母細胞減數分裂的前期,例如多種直翅目和雙翅目昆蟲,部分兩棲類以及獼猴和人等哺乳動物。線蟲的燈刷染色體可出現在間期體細胞中。在植物界,某些大型單細胞藻類含有燈刷染色體。高等植物雄性配子減數分裂中也出現燈刷染色體,例如垂花蔥和玉米。
不同物種燈刷染色體的大小十分懸殊。一般說來,凡單套染色體中DNA的含量(C值)高的動物,燈刷染色體大,側環長,而且燈刷染色體持續時間也比較長。兩棲類的具有最高的C值,燈刷染色體也最大,是研究燈刷染色體的常用材料,其中研究得最為詳盡的是某些有尾兩棲類。
兩棲類燈刷染色體 兩棲類卵母細胞中,第一次減數分裂的雙線期時,每條染色體的兩條同源染色體經複製而形成總數為4條姊妹染色單體的結構並產生交叉(圖1)。這一狀態在卵母細胞中可維持數月或數年之久。
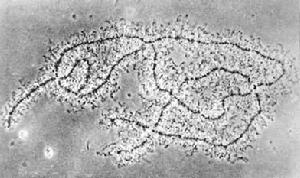 燈刷染色體
燈刷染色體 燈刷染色體
燈刷染色體著絲粒 燈刷染色體的著絲粒通常位於交叉附近。著絲粒有二種形態:一種為顆粒狀,大小形態與前後染色粒不易區分,如東方蠑螈;另一種在著絲粒的前後帶有由相鄰染色粒彼此融合而成的軸棒,例如冠螈(圖3)。著絲粒和軸棒上均無側環。燈刷染色體著絲粒指數與體細胞的大抵相同。
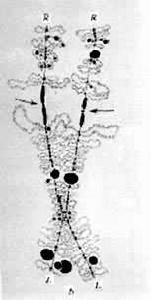 燈刷染色體
燈刷染色體 燈刷染色體
燈刷染色體 燈刷染色體
燈刷染色體球體 在特定部位出現的球體(圖6)相當於一般染色體的次縊痕,並含有酸性蛋白。球體周期性地脫落到核質中,並在同一部位形成新的球體。球體間常見彼此融合現象,但大小並不增加。電子顯微鏡下可見球體內層為圓而緻密的髓心被電子密度較弱的外鞘包圍著(圖7)。 在不同的亞種之間球體的大小是相對恆定的。
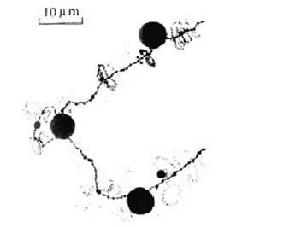 燈刷染色體
燈刷染色體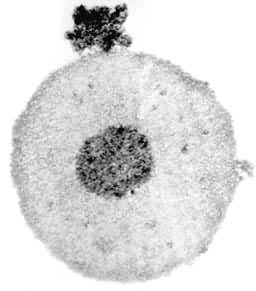 燈刷染色體
燈刷染色體軸顆粒 結構與端粒相似。也可見到軸顆粒間的融合。有些軸顆粒在染色體上具明顯而較恆定的位置,可作為鑑定染色體的界標。
其他界標和工作圖譜 在某些兩棲類燈刷染色體軸上尚有其他界標,如雙軸結構、軸棒狀結構,幾對較大的側環成簇地出現在軸的特定部位等。在形態學基礎上現在已經繪出了多種兩棲類燈刷染色體的工作圖譜(圖8)。這樣的圖譜反映一整套燈刷染色體的相對長度、著絲粒部位和各種不同界標著生的位置,是利用燈刷染色體進行深入研究的基礎。
 燈刷染色體
燈刷染色體果蠅精母細胞第一次成熟分裂到雙線期,在某一染色體上會出現成對的側環。由於XO型的果蠅的精母細胞中不具側環,而XYY果蠅的精母細胞中側環數目加倍, 因而很容易判定只有Y染色體是燈刷染色體。Y染色體在減數分裂中不配對,所以和兩棲類的不同,果蠅的燈刷染色體不是二價體。果蠅的 Y染色體共有平均長度為50微米 的5對側環(圖9),每對側環都有專門的名稱。
 燈刷染色體
燈刷染色體杵狀環 位於長臂近著絲粒處,帶有2個體積很大的杵狀體故名。杵狀體由0.2~1微米的蛋白質球體和彌散的纖絲物質組成。這對側環是否象燈刷側環那樣正在轉錄,目前還缺乏染色質鋪展實驗的證據。
管狀帶 這對側環在光學顯微鏡可見由含緻密小顆粒的彌散物質組成。在電子顯微鏡下可見彌散物質由直徑35納米的小管組成,而緻密的顆粒由小管緊密包裝而成。鋪展實驗表明管狀帶的結構類似燈刷側環。
假核仁 這對側環具有大的假核仁和前後各一個假核仁球部,兩者超微結構相似,均由纖絲物質組成,其中含有小的 RNP顆粒。鋪展實驗中可見轉錄本長達1000kb,說明這對側環上可能只有一個轉錄單位。
絲狀體 位於長臂端部,由兩部分組成。遠端的彌散部分由纖絲組成,其中分布著35~40納米的富含RNP的顆粒,一端與假核仁球部相接;近端的緻密部分由緊密包裝的纖絲組成,無RNP顆粒。鋪展實驗證明DNP軸上有長約500kb的轉錄單位。
果蠅燈刷染色體的側環活躍地合成其他染色體不能合成的一種對生育必不可少的RNA。沒有Y染色體的XO型雄果蠅是完全不育的。 Y染色體有缺陷的果蠅精子發生中出現的形態學異常是十分多樣的:精子細胞在伸長時生長受阻;精子中細胞器格局被打亂以及這些細胞器出現種種異常。這就說明側環上的基因的機能(即Y染色體雄性生育因子)不是為這些細胞器分化時所需的某一蛋白質編碼,而是調節著細胞生長和精子細胞器的自我裝配。而且這種Y染色體畸形的果蠅,其精子發生受阻在早期或晚期,取決於缺少哪一對側環。這提示不同的生育因子可能依次地作用於精子形成的不同的階段。
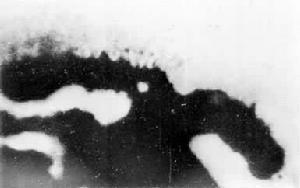
植物的燈刷染色體 有關植物燈刷染色體的報導不多。在垂花蔥雄性減數分裂雙線期、玉米雄性減數分裂終變期均有燈刷期的記載。但所報導的燈刷染色體,只是一條較長的染色體,周圍有絨毛狀的結構(圖10)。真菌減數分裂雙線期的燈刷染色體與此大同小異。只有地中海傘藻具有典型的燈刷染色體的結構(圖11)。
 燈刷染色體
燈刷染色體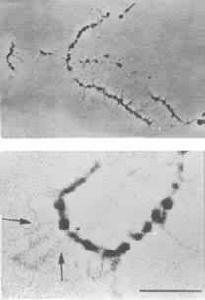 燈刷染色體
燈刷染色體70年代以來,曾利用同位素標記的DNA與燈刷染色體上新產生的 RNA進行原位雜交,以確定單個基因的定位。但是陸續發現這些DNA探針專一性不強,例如組蛋白基因和5sDNA探針都能與許多側環上轉錄的RNA雜交,而燈刷染色體同一區域也可為不同探針所標記,這樣就使人們對用這種方法所定位的基因的真實性產生了懷疑。
隨著更精確的實驗技術的發展和套用,可以預期在卵子發育中燈刷染色體的基因活動將逐漸得到闡明,這不僅對於理解胚胎早期發育十分重要,而且還可以幫助了解體細胞分化中基因的調節與控制。
