概述
雌雄同株植物中,雄蕊發育不正常,不能產生有功能的花粉,但它的雌蕊發育正常,能接受正常花粉而受精結實,並能將雄性不育性遺傳給後代的植物品系。
雄性不育一般可分為3種類型。(1)細胞質雄性不育型,簡稱質不育型,表現為細胞質遺傳。通常以單一的細胞質基因S和N分別代表雄性不育和雄性可育。用可育株花粉給雄性不育株雌蕊授粉,能正常結實,但F1植株仍表現為雄性不育的母體性狀,因而不能自交產生F2,農業生產上不能利用。(2)細胞核雄性不育型,簡稱核不育型,表現為細胞核遺傳,雄性不育性大多為一對隱性基因(msms)所控制,正常可育性為相對的顯性基因(MsMs)所控制。雄性不育株與正常株雜交,F1植株為雄性可育(MsMs);F1自交產生的F2,可育株與不育株之比為3∶1,難以用普通方法保持雄性不育系,在農業生產上也不可能廣泛利用。(3)核-質互作不育型,表現為核-質互作遺傳。不但需要細胞質有不育基因S,而且需要細胞核里有純合的不育基因(rfrf),二者同時存在,方能使植株表現為雄性不育。
如胞質基因為可育N,則不論核基因是可育(RfRf)還是不育(rfrf),都表現為雄性可育。同樣,如核里具有可育基因(RfRf)或(Rfrf),則不論胞質基因是可育N還是不育S,也都表現為雄性可育。這種由核-質互作形成的雄性不育系,其遺傳組成為S(rfrf),不能產生正常的花粉,但可作為雜交母本。由於能找到保持系N(rfrf)[用它與不育系雜交,所產生的F1仍能保持雄性不育,即:S(rfrf)(♀)×N(rfrf)(♀)→S(rfrf)(F1)(不育)]並能接受恢復系S(RfRf)或N(RfRf)[用它們與不育系雜交,所產生的F1都是可育的,即:S(rfrf)(♀)×S(RfRf)→S(Rfrf)(F1)(可育),或S(rfrf)(♀)×N(RfRf)→S(Rfrf)(F1)(可育)]的花粉,使F1恢復為雄性可育,F1植株自交產生F2,所以在農業生產上可以廣泛套用。雄性不育系可以免除人工去雄,節約人力,降低種子成本,還可保證種子的純度。目前水稻、玉米、高粱、洋蔥、蓖麻、甜菜和油菜等作物已經利用雄性不育系進行雜交種子的生產;對其他作物的雄性不育系,也正在進行廣泛的研究。
細胞核雄性不育系
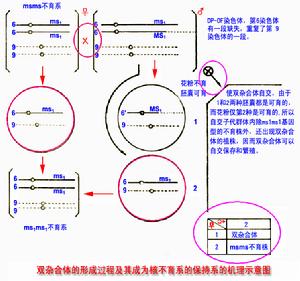 原理圖
原理圖即由控制花粉正常育性的核基因發生突變而形成的不育系。
不育機制
一般由1對隱性基因控制,但也有由2~3對隱性基因互作而產生的雄性不育性(如萵苣)。假如控制花粉正常育性是一對顯性基因RfRf,則由於隱性突變,雜合體Rfrf自交後將會分離出純合基因型rfrf,表現為雄性不育。大麥、玉米、高粱、大豆、番茄、棉花等很多作物都有這樣的突變體。但偶爾還發現有雜合的顯性核不育現象。其正常可育的基因型為msms,而經顯性突變後產生的雜合基因型Msms會由於Ms的顯性作用表現為雄性不育,當它被正常育性植株msms授粉結實時,其子代按1:1比例分離出顯性不育株和隱性可育株,並依此方法代代相傳。1972年中國在山西省發現的由顯性單基因控制的太谷核不育小麥就屬於此類。
利用
因隱性核不育系難以找到有效的保持系,故不能大量產生不育系種子供制種用;但可用雜合可育株給不育株授粉,在正常育性受1對顯性基因控制的情況下,其子代將按1:1比例分離出純合不育株和雜合可育株。用雜合可育株對不育株授粉,下一代育性分離仍是1:1的比例。採用這種作法可以較大量地繁殖不育株與可育株的混合群體。這種群體內既有不育株又有保持不育性能力的植株,有人因此稱之為兩用系。雜交制種時,必須在開花前剔去母本群體內的可育株,以保證制種的純度。一般栽培品種都可作隱性核不育系的恢復系,因此易於配出強優勢組合。但要在混合群體開花前的短促時間內剔除全部可育株,對於繁殖係數低、用種量大的作物常因十分費工而不易做到。1965年,美國R.T.拉梅奇為解決大麥核不育系種子繁殖的困難,提出利用“平衡三級三體”的遺傳機制:即在正常染色體上具有隱性雄性不育和隱性稃色正常的基因,在額外染色體上有相應的顯性可育基因,並在其附近設法引入一個能使稃殼有色的顯性標誌基因,兩者緊密連鎖。額外染色體一般不能由花粉傳遞,只能以30%的比例由雌配子傳給下代。這樣的三級三體自交後將產生二體和三體兩類植株,二體植株具純合的雄性不育基因和正常稃色;三體植株帶有一個顯性可育基因和有色稃殼。通過光電比色裝置對種子稃色進行篩選,可將帶雄性可育基因的有色種子剔除,以繁殖純不育系。這一構想後得到實現,育成了1個大麥雜交種,並在生產上推廣。但後來在推廣繁殖過程中,發現額外染色體通過雄配子的傳遞率比預期的高,上述機制受到干擾,而且雜種優勢不夠強,因而停止套用。對於繁殖係數高、用種量少的作物如番茄等,則可直接套用兩用系作母本,於開花前逐株檢查育性並剔除可育株,授以父本恢復系花粉,產生雜交種子。總之,核不育系由於難以找到保持系,目前在生產上仍不能有效利用。而單基因控制的太谷顯性核不育小麥在沒有作出標誌基因之前,只能作為常規育種中開展輪迴選擇和回交育種的親本之用。
質、核互作或細胞質雄性不育系
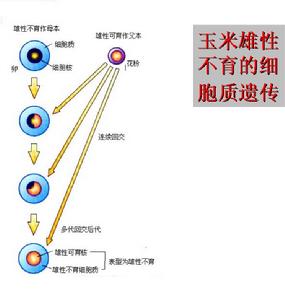 玉米雄性不育細胞質遺傳
玉米雄性不育細胞質遺傳即由細胞質基因與核基因互作而產生的雄性不育系。
不育機制
若胞質記憶體在不育基因S,而在相應的細胞核內又具有1對純合隱性不育基因rfrf,則這種具有S(rfrf)基因型的植株就表現為雄性不育。但若核內為純合或雜合的顯性恢復基因即RfRf或Rfrf所代替,胞質S基因的作用便受到抑制而使植株表現為可育。如果胞質記憶體在著可育基因N,則不論核基因是否可育,其植株全都表現可育。可見對這類不育性起主導作用的是胞質基因。胞質基因S可能來自突變,也可通過核置換將栽培品種的核通過多次回交法導入遠緣的屬、種、亞種以及地理遠緣品種的胞質中,利用核質間不協調而產生雄性不育性。如水稻野敗型、高粱邁羅型、玉米T型和C型、小麥提莫菲維型和擬斯卑爾脫型等雄性不育系都屬於這一類。
利用
質、核互作雄性不育系的優點是易於三系配套。基因型為N【rfrf】的一般栽培品種都可成為保持系,而基因型為S【RfRf】或N【RfRf】的栽培品種或雜交選系可成為恢復系。1970年中國在海南島崖縣發現野敗型水稻不育株,經選用栽培稻品種連續回交數代,育成了野敗型不育系,上述輪迴親本便分別成為相應的保持系。此後又經測交篩選,發現IR24等引入品種對野敗型不育系具有很強的恢復力。此外,還用雜交導入恢復基因等方法選育出一批優良恢復系。質、核互作不育系在生產利用上比較簡便,只要設定兩個隔離區,即不育系繁殖區和制種隔離區,就可以生產大量雜交種。現已廣泛套用於水稻、高粱、玉米、甜菜、洋蔥、蠟燭稗等作物,收到了很大經濟效益。但也發現某些不育胞質會帶來一些不利影響。例如,玉米T型不育胞質是小斑病菌T小種專化侵染的對象,美國一度因此受到較大的經濟損失;T型不育胞質使小麥雜交當代種子皺癟和發芽率降低等。但隨著育種研究的進展,這些缺點正在得到克服。質、核互作不育系作為雜種優勢利用的有效手段正在日益發揮著重要的作用。
