
漢語詞語
基本信息
【詞目】擴散【拼音】kuò sàn【詞性】動詞【解釋】擴大分散出去。詳細解釋
擴散可以分類為很多不同種類的擴散,其需要和狀態大體不相同。有些擴散需要介質,而有些則需要能量。因此不能將不同種類的擴散一概而論。【示例】化學名詞
概念
擴散(diffusion):物質分子從高濃度區域向低濃度區域轉移,直到均勻分布的現象。擴散的速率與物質的濃度梯度成正比。由於分子(原子等)的熱運動而產生的物質遷移現象.一般可發生在一種或幾種物質於同一物態或不同物態之間,由不同區域之間的濃度差或溫度差所引起,前者居多.一般從濃度較高的區域向較低的區域進行擴散,直到同一物態內各部分各種物質的濃度達到均勻或兩種物態間各種物質的濃度達到平衡為止.顯然,由於分子的熱運動,這種“均勻”、“平衡”都屬於“動態平衡”,即在同一時間內,界面兩側交換的粒子數相等,如紅棕色的二氧化氮氣在靜止的空氣中的散播,藍色的硫酸銅溶液與靜止的水相互滲入,鋼製零件表面的滲碳以及使純淨半導體材料成為N型或P型半導體摻雜工藝等等都是擴散現象的具體體現;在電學中半導體PN結的形成過程中,自由電子和空穴的擴散運動是基本依據.擴散速度在氣體中最大,液體中其次,固體中最小,而且濃度差越大、溫度越高、參與的粒子質量越小,擴散速度也越大。擴散過程,是分子掙脫彼此間分子引力的過程,這個過程,分子需要能量來轉化為動能,也就需要從外界吸收熱量。晶體學中,擴散是物質內質點運動的基本方式,當溫度高於絕對零度時,任何物系內的質點都在作熱運動。當物質內有梯度(化學位、濃度、應力梯度等)存在時,由於熱運動而導致質點定向遷移即所謂的擴散。因此,擴散是一種傳質過程,巨觀上表現出物質的定向遷移。在氣體和液體中,物質的傳遞方式除擴散外還可以通過對流等方式進行;在固體中,擴散往往是物質傳遞的唯一方式。擴散的本質是質點的無規則運動。晶體中缺陷的產生與複合就是一種巨觀上無質點定向遷移的無序擴散。晶體結構的主要特徵是其原子或離子的規則排列。然而實際晶體中原子或離子的排列總是或多或少地偏離了嚴格的周期性。在熱起伏的過程中,晶體的某些原子或離子由於振動劇烈而脫離格點進入晶格中的間隙位置或晶體表面,同時在晶體內部留下空位。顯然,這些處於間隙位置上的原子或原格點上留下來的空位並不會永久固定下來,它們將可以從熱漲落的過程中重新獲取能量,在晶體結構中不斷地改變位置而出現由一處向另一處的無規則遷移運動。在日常生活和生產過程中遇到的大氣污染、液體滲漏、氧氣罐泄漏等現象,則是有梯度存在情況下,氣體在氣體介質、液體在固體介質中以及氣體在固體介質中的定向遷移即擴散過程。由此可見,擴散現象是普遍存在的。晶體中原子或離子的擴散是固態傳質和反應的基礎。無機材料製備和使用中很多重要的物理化學過程,如半導體的摻雜、固溶體的形成、金屬材料的塗搪或與陶瓷和玻璃材料的封接、耐火材料的侵蝕等都與擴散密切相關,受到擴散過程的控制。通過擴散的研究可以對這些過程進行定量或半定量的計算以及理論分析。無機材料的高溫動力學過程——相變、固相反應、燒結等進行的速度與進程亦取決於擴散進行的快慢。並且,無機材料的很多性質,如導電性、導熱性等亦直接取決於微觀帶電粒子或載流子在外場——電場或溫度場作用下的遷移行為。因此,研究擴散現象及擴散動力學規律,不僅可以從理論上了解和分析固體的結構、原子的結合狀態以及固態相變的機理;而且可以對無機材料製備、加工及套用中的許多動力學過程進行有效控制,具有重要的理論及實際意義。過程
擴散[1] (dispersal),生物個體或其傳布體(如孢子、種子)向其他地域傳布的過程。又稱散布。各種生物都能通過不同的擴散方式擴大棲居範圍和分布區域;擴散還促進生物群落的演替,增加物種的多樣性;大範圍的擴散結合地理隔離是物種分化的重要條件;新的環境意味著不同的自然選擇機制,可改變種群的基因頻率,因此擴散是促進生物進化的重要因素。現象
生物從一地區傳布至另一地區是常見的自然現象,這種異地間的物種交流,促進了各地生物群落的演替,增加了各地區物種的多樣性。大範圍的擴散結合地理隔離是物種分化的重要條件;對物種來說,新的環境意味著不同的自然選擇機制,它將改變種群的基因頻率,因此擴散是促進生物種系進化的重要因素之一。火山爆發毀滅原地生物後,風和洋流會帶來新的生命,各種微生物、植物和動物相繼在此定居。先擴散來的是對異常環境耐受力高的先驅物種,繼之是在安定環境裡競爭能力強的物種,它們在前者開拓的基礎上建立起穩定的群落。
類型
主動自力生物擴散包括主動(自力)和被動(借外力)兩類。缺乏行動能力的微生物和植物大多靠被動傳布〔見傳播(植物)〕,動物則多行主動擴散。主動擴散 生物的生存繁衍需要空間、食物和配偶等條件。生物個體過於稠密時,對空間的需要不易滿足,食源不足及環境惡化會限制種群的發展。與植物相比,動物具有主動尋找生存空間、食源和躲避危險的能力。動物的主動擴散多發生在生殖期。許多鳥、獸在交配季節四處尋找交配和生育的場所,當其出生地附近的空間多被占據時,就被迫遠距離擴散。其中多數個體往往會因遇到不利環境而死亡,但只要少數個體遇到食源豐富且天敵稀少的合適生境,便可能建立起新的種群,使種的分布區擴大。表現
許多昆蟲也有很大的擴散能力,而且有些種還表現出多態現象,即同一物種中存在兩種或多種形態和行為均不相同的類型,其中之一專營擴散,具翅或具其他有利於擴散的結構。有的多態現象為遺傳性的,如灰線小卷蛾有兩類不同的遺傳型,一類的幼蟲不耐密集而易擴散,另一類則否;兩者共存互不替代,但數量比例隨種群密度而變。非遺傳性的多態現象則有不同的基礎,有的是發育性的,如蝴蝶、蜻蜓及蜂類的無翅幼蟲取食而不擴散,成蟲則擴散而極少取食。有些昆蟲如蜉蝣的成蟲,甚至完全不取食,僅擴散、交尾、產卵直至死亡。蟻和白蟻的情況也類似。這類社群性昆蟲中,不同社群成員(不同階級)的結構與行為特化,各執行不同的職能,僅有翅型負責擴散到新區域尋找新的繁殖基地。這兩類事例都屬於常規的分工現象,一個是幼蟲與成蟲間的分工,另一個是社群成員間的分工。但飛蝗的多態現象則屬非常規性的,只有當種群密度增加、食物缺乏時,才由散居型經過型變轉化為群居型(即擴散型),造成大規模的擴散,所至之處穀草被其掠食一空。植物中也有主動擴散的事例,如許多豆科植物果實成熟時果莢裂開將種子散落出去;鳳仙花的果實成熟後,若受外力觸碰,果皮即向內捲縮將種子彈出。不過這種擴散的能力不高,傳播範圍有一定局限。被動擴散 主要依賴氣流(風)、水流或其他生物的攜帶。風力傳布 微生物和微小的孢子、種子很容易由風傳送,甚至一些較大的動物也能被大風卷帶很遠。在大洋中部的上空,曾收集到昆蟲和蜘蛛。標記重捕實驗證實,粘蟲可隨3、4月份在中國南方盛行的西南氣流成群遷飛,從華南諸省擴散到江淮流域。還證實,每年6月在日本稻田中褐飛虱數量突增,它們是從中國隨風傳入的。無翅幼蟲的擴散常以風力傳送為主。蓑蛾幼蟲在母蟲護囊內孵化之後,成群從囊下排泄孔鑽出,吐絲下垂,隨風向四周擴散。有的植物的種子小如粉塵(如蘭科植物),有的植物種子具翅(槭、榆等)、毛(柳、蒲公英等)和氣球狀囊(如酸漿)等特化結構,這都有利於隨風擴散。水流傳布 借水流擴散的陸地生物必須能浮在水上,不易被水浸透,並能保持內部的滲透平衡。有些專借水流擴散的植物果實表面具不易透水的蠟質或纖維,其組織內含氣,故體輕而不沉。椰子的表面具發達的纖維層,能在水面長期漂流。巨大的洋流都有固定的行程,因此河口或淡水的水生動物,甚至某些無飛翔能力的陸生動物都可能隨洋流由一個大陸擴散至另一個大陸。有些動物雖會游泳,但其遠距離擴散則可能是隨漂木、浮冰實現的。動物傳布 寄生生物是借寄主擴散的,有的寄生生物在其生活史的各個階段寄生於不同的寄主。轉換寄主的過程或為主動或為被動。有的寄生物(如血吸蟲)借本身的行動尋找下一個寄主,而有的(如瘧原蟲)則完全被動地擴散,蚊蟲吸食不同寄主個體的血液時瘧原蟲隨之侵染新的寄主。很多生物依附在其他生物的體表擴散。依附的方式很多:有些植物的果實和種子具刺或鉤(如蒼耳的果實),還有的具粘性分泌物(如亞麻種子);一些小昆蟲,蟎類等常粘附在脊椎動物的皮毛上,也有些小動物粘附在候鳥的足上。有的植物種子經動物取食而擴散。例如草食動物經常吃下大量種子,這些種子可隨糞便排至多處。很多肉質果實憑其色、香、味吸引動物取食,其種子或被吐出或隨糞便排出。這些種子一般均能抵抗動物消化液的侵蝕,有些種子只有在種皮經消化道作用後才能萌發。人類也是重要的生物擴散因子。長途的人貨流通有意無意地擴散了大量生物。棉花的主要害蟲棉紅鈴蟲原產印度,20世紀30年代傳入中國沿海地區,現在除新疆外已遍及中國各棉區。此外,人類還有意識地引種或培育很多優良的穀物、林木和家禽家畜,並在全世界範圍推廣。擴散力的進化 在自然界,擴散力存在兩種截然不同的進化趨勢,一種是擴散力增強,另一種是減弱甚或消失。例如加拿大不列顛哥倫比亞省省會維多利亞附近的溫哥華島的西部天幕毛蟲,在其種群中,擴散力弱的個體在生境附近產卵,後代常死於惡劣氣候;擴散力強的個體向遠處產卵,生於較有利環境中的後代可能存活,並且大多是活潑個體。如此代代選擇,該種的擴散能力不斷增強。另一方面,如果靠擴散找到未開拓的適宜環境的機會很小時,自然選擇將對抗擴散。例如在海島和山巔環境中不會飛的鳥和昆蟲種數比平坦的大陸上要多。達爾文在考察馬德拉群島時注意到,島上550種甲蟲中有200種有翅卻缺乏飛翔能力,而當地29個屬甲蟲中有23個屬於狂風時潛藏,天氣較好時才外出活動。達爾文認為,這種情況主要是因為善飛個體在飛行中可能被狂風吹入大海,而翅發育不全或習性怠惰的個體,反能更多地存留下來。動物地理學家P.J.達林頓認為,遇敵時翅膀有助於逃生,但海島上天敵很少或沒有,翅就成為無用之物。與此類似,島上植物的擴散力也明顯下降或幾乎完全消失。有些植物果實變大,但用於傳布的附器(如倒鉤)卻未相應增加,有的種子附器甚至退化,這類種子或果實的數量也常相應地減少。這種進化變異可減少向島外的無效擴散,而個體增大則可更多地儲存營養,反而有利於幼苗生長。人工選擇也可改變擴散能力。例如人類栽培農作物要求的是高收穫量,而不是高擴散力。目前種植的小麥、水稻等穀物的種子散落性已消失,亞麻和罌粟蒴果的開裂機制也已退化。擴散現象的數學模型 根據觀察種子散落的情況和用標記重捕法進行的昆蟲擴散試驗,人們發現,隨著擴散距離的增加,可能找到的傳布體的數量按指數比率下降。種子散落屬於被動擴散;有些小昆蟲飛翔力不強,方向和距離也基本是隨機的,所以也表現上述規律。擴散是研究種群數量變動的重要因子。在生產實踐中,害蟲和雜草的擴散會給人類健康和農牧業帶來損害。因此,了解生物擴散的特性,對防治有害生物有重要意義。許多細菌性和病毒性人畜疾病及作物病害的防治,往往取決於切斷病原體的傳布途徑。稻縱卷葉螟和褐飛虱是中國南方成群長距離擴散的重要水稻害蟲,田間種群數量經常突增或突減,掌握其擴散規律便可預測它的發生量和發生期。利用捕食性天敵防治害蟲時,必須研究天敵和害蟲在各種環境(包括不同的地形、植物長勢、氣候變化等)中的擴散能力、傳布方向和可能的傳布距離等,以便確定人工釋放天敵的數量、野外釋放點布局和釋放點高度等。物理學名詞
指一種物質的分子分散到另一種物質的分子中,最後均勻分布管理學/傳播學名詞
擴散(diffusion)是創新通過一段時間,經由特定的渠道,在某一社會團體的成員中傳播。它是特殊類型的傳播,所含信息與新觀念有關。擴散是一個特殊類型的傳播。傳播的信息是有關一個新的觀念,而觀念之新奇度賦予擴散一種特製。新意味著擴散中含有某種程度上的不確定因素。擴散是一種社會變化,可以被定義為社會系統的結構和功能發生變化的過程。它既包含了自發的傳播,又包含了自覺的傳播,動力氣象學
| ▪動力氣象學 | ▪大氣熱力學 | ▪力能學 | ▪氣體常數 | ▪感熱 |
| ▪可逆絕熱過程 | ▪潛熱 | ▪道爾頓定律 | ▪熱功當量 | ▪不可逆絕熱過程 |
| ▪逆濕 | ▪熱力學方程 | ▪外強迫 | ▪理想流體 | ▪理想氣體 |
| ▪不可壓縮流體 | ▪融[化]點 | ▪逆溫 | ▪湍流逆溫 | ▪廓線 |
| ▪地面逆溫 | ▪輻射逆溫 | ▪輻射冷卻 | ▪覆蓋逆溫 | ▪下沉逆溫 |
| ▪鋒面逆溫 | ▪信風逆溫 | ▪逆溫層 | ▪熱力學圖 | ▪熵 |
| ▪等熵面圖 | ▪等熵分析 | ▪溫熵圖 | ▪焓 | ▪溫度-對數壓力圖 |
| ▪溫度梯度 | ▪溫度平流 | ▪氣塊 | ▪絕熱上升 | ▪絕熱下沉 |
其他科技名詞
| ▪絕熱冷卻 | ▪絕熱增溫 | ▪假絕熱過程 | ▪非絕熱過程 | ▪絕熱過程 |
| ▪對流凝結高度 | ▪等熵凝結高度 | ▪混合凝結高度 | ▪氣溫直減率 | ▪絕熱直減率 |
| ▪乾絕熱直減率 | ▪環境直減率 | ▪超絕熱直減率 | ▪濕絕熱直減率 | ▪凝結高度 |
| ▪湍流凝結高度 | ▪凝結效率 | ▪熱力層結 | ▪層結曲線 | ▪相當溫度 |
| ▪相 | ▪相變 | ▪相函式 | ▪位相譜 | ▪相速[度] |
| ▪平面波 | ▪虛溫 | ▪乾絕熱過程 | ▪絕熱檢驗 | ▪濕絕熱過程 |
| ▪絕熱圖 | ▪假絕熱圖 | ▪假絕熱直減率 | ▪位溫 | ▪假相當位溫 |
| ▪濕球位溫 | ▪假濕球位溫 | ▪假濕球溫度 | ▪熱力學霜點溫度 | ▪熱力學冰球溫度 |
| ▪熱力學濕球溫度 | ▪大氣動力學 | ▪空氣動力學 | ▪運動方程 | ▪連續方程 |
| ▪不連續[性] | ▪狀態方程 | ▪波爾茲曼常數 | ▪普適氣體常數 | ▪閉合系統 |
| ▪能量守恆 | ▪質量守恆 | ▪保守性 | ▪渦度方程 | ▪正壓渦度方程 |
| ▪散度方程 | ▪流體靜力方程 | ▪布西內斯克近似 | ▪布西內斯克方程 | ▪流體靜力近似 |
| ▪ω方程 | ▪擾動方程 | ▪熱成風方程 | ▪水汽[守恆]方程 | ▪平流方程 |
| ▪馬古列斯公式 | ▪無量綱方程 | ▪平衡方程 | ▪無量綱參數 | ▪因子分析 |
| ▪斜壓過程 | ▪雷諾應力 | ▪非黏性流體 | ▪氣象風洞 | ▪相當正壓大氣 |
| ▪靜力適應過程 | ▪半地轉理論 | ▪地轉適應 | ▪半地轉方程 | ▪有效位能 |
| ▪位能 | ▪渦動擬能 | ▪位勢米 | ▪動力米 | ▪位勢高度 |
| ▪動力高度 | ▪聲重力波 | ▪重力流 | ▪慣性不穩定 | ▪靜力不穩定[性] |
| ▪自由對流高度 | ▪抬升凝結高度 | ▪抬升指數 | ▪不穩定空氣 | ▪不穩定氣團 |
| ▪[大氣]穩定度 | ▪[大氣]不穩定度 | ▪靜力穩定度 | ▪穩定度指數 | ▪流體靜力不穩定度 |
| ▪絕對穩定 | ▪絕對不穩定 | ▪中性穩定 | ▪條件[性]不穩定 | ▪慣性穩定度 |
| ▪對流不穩定 | ▪對流[性]穩定度 | ▪動力穩定[性] | ▪動力不穩定[性] | ▪穩定度參數 |
| ▪穩定空氣 | ▪巨觀黏滯度 | ▪大氣擾動 | ▪吸引子 | ▪奇怪吸引子 |
| ▪分岔 | ▪有限振幅 | ▪自由波 | ▪角展寬 | ▪非線性不穩定[性] |
| ▪薄片法 | ▪第二類條件[性]不穩定 | ▪切變不穩定 | ▪切變層 | ▪正壓不穩定 |
| ▪斜壓不穩定 | ▪潛在不穩定 | ▪位渦守恆 | ▪傾向方程 | ▪z坐標 |
| ▪p坐標 | ▪σ坐標 | ▪自然坐標[系] | ▪位溫坐標 | ▪混合坐標 |
| ▪歐拉坐標 | ▪氣壓坐標系 | ▪曲線坐標 | ▪柱面坐標 | ▪天赤道 |
| ▪天球 | ▪天極 | ▪寒極 | ▪冷池 | ▪冰凍學 |
| ▪赤緯 | ▪赤經 | ▪黃道 | ▪磁傾角 | ▪磁傾赤道 |
| ▪局地軸 | ▪輻合 | ▪匯流 | ▪分流 | ▪摩擦輻合 |
| ▪摩擦曳力 | ▪輻散 | ▪摩擦輻散 | ▪散度 | ▪微分分析 |
| ▪散度定理 | ▪局部導數 | ▪水平散度 | ▪引導氣流 | ▪引導高度 |
| ▪渦度 | ▪正渦度平流 | ▪相對渦度 | ▪絕對渦度 | ▪行星渦度 |
| ▪行星反照率 | ▪行星渦度效應 | ▪渦度平流 | ▪β平面 | ▪絕對渦度守恆 |
| ▪地轉渦度 | ▪地轉平流 | ▪點渦 | ▪熱渦度 | ▪氣鏇性渦度 |
| ▪反氣鏇[性]渦度 | ▪切變渦度 | ▪曲率渦度 | ▪位勢渦度 | ▪渦動傳導率 |
| ▪渦動黏滯率 | ▪動力黏性 | ▪摩擦速度 | ▪渦動切應力 | ▪卡門常數 |
| ▪大氣波動 | ▪強迫振盪 | ▪群速度 | ▪共振理論 | ▪長波調整 |
| ▪慣性波 | ▪慣性圓 | ▪慣性力 | ▪慣性振盪 | ▪斜壓波 |
| ▪斜壓性 | ▪正壓波 | ▪正壓性 | ▪重力波 | ▪慣性重力波 |
| ▪內波 | ▪重力內波 | ▪重力外波 | ▪縱波 | ▪重力波拖曳 |
| ▪卡爾曼濾波 | ▪拖曳係數 | ▪地形拖曳 | ▪地形波 | ▪背風波 |
| ▪亥姆霍茲波 | ▪亥姆霍茲不穩定 | ▪開爾文波 | ▪開爾文-亥姆霍茲波 | ▪蘭姆波 |
| ▪羅斯貝波 | ▪定常波 | ▪瞬變波 | ▪孤立波 | ▪退行 |
| ▪前進波 | ▪後退波 | ▪切變波 | ▪駐波 | ▪混合羅斯貝重力波 |
| ▪超長波 | ▪不穩定波 | ▪大氣噪聲 | ▪普朗特數 | ▪瑞利數 |
| ▪雷諾數 | ▪里查森數 | ▪整體裡查森數 | ▪臨界裡查森數 | ▪通量里查森數 |
| ▪布倫特-韋伊塞萊頻率 | ▪伯格數 | ▪弗勞德數 | ▪羅斯貝數 | ▪羅斯貝參數 |
| ▪羅斯貝圖解 | ▪馬赫數 | ▪轉盤實驗 | ▪哈得來域 | ▪羅斯貝域 |
| ▪白貝羅定律 | ▪浮力 | ▪浮力速度 | ▪整體平均 | ▪徑向流入 |
| ▪混合層 | ▪無輻散層 | ▪鏇衡風 | ▪梯度風 | ▪反梯度風 |
| ▪次梯度風 | ▪超梯度風 | ▪熱成風 | ▪順轉風 | ▪逆轉風 |
| ▪風切變 | ▪徑向風 | ▪地轉風 | ▪摩擦風 | ▪次地轉風 |
| ▪超地轉風 | ▪非地轉風 | ▪變壓風 | ▪角動量 | ▪角動量平衡 |
| ▪基本氣流 | ▪急流 | ▪急流核 | ▪極夜急流 | ▪北支急流 |
| ▪南支急流 | ▪夜間急流 | ▪副熱帶急流 | ▪急流軸 | ▪流線 |
| ▪西風急流 | ▪對流雲 | ▪深對流 | ▪穿透對流 | ▪淺對流 |
| ▪線對流 | ▪深水波 | ▪淺水波 | ▪埃克曼流 | ▪埃克曼抽吸 |
| ▪熱對流 | ▪熱泡 | ▪濕對流 | ▪乾對流 | ▪積雲對流 |
| ▪夾卷 | ▪夾卷率 | ▪中尺度對流系統 | ▪暖平流 | ▪冷平流 |
| ▪上升氣流 | ▪下沉氣流 | ▪對流調整 | ▪參數化 | ▪對流參數化 |
| ▪淺對流參數化 | ▪中尺度對流複合體 | ▪地轉運動 | ▪絕對角動量 | ▪角動量守恆 |
| ▪準地轉運動 | ▪半地轉運動 | ▪非地轉運動 | ▪無鏇運動 | ▪無輻散運動 |
| ▪準無輻散 | ▪斜壓大氣 | ▪正壓大氣 | ▪模態 | ▪斜壓模[態] |
| ▪正壓模[態] | ▪正規模[態] | ▪埃克曼層 | ▪埃克曼螺線 | ▪風速對數廓線 |
| ▪大氣環流 | ▪主級環流 | ▪三級環流 | ▪經向環流 | ▪緯向環流 |
| ▪哈得來環流[圈] | ▪沃克環流[圈] | ▪直接環流[圈] | ▪間接環流[圈] | ▪信風環流 |
| ▪環流圈 | ▪局地環流 | ▪城鄉環流 | ▪三圈[經向]環流 | ▪費雷爾環流 |
| ▪環流指數 | ▪高指數 | ▪低指數 | ▪指數循環 | ▪環流定理 |
| ▪開爾文環流定理 | ▪伯努利定理 | ▪調和函式 | ▪皮葉克尼斯環流定理 | ▪越赤道氣流 |
| ▪環流調整 | ▪中間層環流 | ▪大尺度環流 | ▪大氣活動中心 | ▪平流層爆發 [性]增溫 |
| ▪平流層耦合作用 | ▪平流層引導 | ▪大氣湍流 | ▪湍流相似理論 | ▪二維湍流 |
| ▪渦鏇 | ▪渦動相關 | ▪渦動平流 | ▪卡門渦街 | ▪瞬變渦動 |
| ▪通量 | ▪湍流通量 | ▪各向同性湍流 | ▪湍流K理論 | ▪普朗特混合長理論 |
| ▪混合長 | ▪運動黏滯性 | ▪湍流邊界層 | ▪層流邊界層 | ▪湍流譜 |
| ▪機械湍流 | ▪湍流能量 | ▪耗散 | ▪耗散率 | ▪擴散 |
| ▪擴散率 | ▪擴散模式 | ▪湍流交換 | ▪湍流擴散 | ▪E-P通量 |
| ▪側向混合 | ▪分子黏性 | ▪分子黏滯係數 | ▪混沌 | ▪數值天氣預報 |
| ▪資料同化 | ▪傅立葉分析 | ▪快速傅立葉變換 | ▪濾波 | ▪細格線 |
| ▪模式輸出統計預報 | ▪相當正壓模式 | ▪準地轉模式 | ▪過濾模式 | ▪絕熱模式 |
| ▪輻射模式 | ▪原始方程模式 | ▪中尺度模式 | ▪半球模式 | ▪消轉時間 |
| ▪起轉時間 | ▪尺度分析 | ▪量綱分析 | ▪天氣尺度 | ▪行星尺度 |
| ▪中尺度 | ▪α中尺度 | ▪β中尺度 | ▪γ中尺度 | ▪蒙特卡羅方法 |
| ▪譜模式 | ▪離散譜 | ▪離散化 | ▪頻散關係 | ▪求積譜 |
| ▪可移動細格線模式 | ▪半隱式格式 | ▪守恆格式 | ▪差分模式 | ▪柯朗-弗里德里希斯-列維條件 |
| ▪迎風差格式 | ▪計算不穩定 | ▪小擾動法 | ▪氣象噪聲 | ▪能量串級 |
| ▪能量密度譜 | ▪客觀分析 | ▪方差分析 | ▪變分客觀分析 | ▪物理模[態] |
| ▪計算模[態] | ▪模式解析度 | ▪技巧[評]分 | ▪次格線尺度過程 | ▪次格線[尺度]參數化 |
| ▪正規模[態]初值化 | ▪初值化 | ▪內積 | ▪動力初值化 | ▪靜力初值化 |
| ▪四維資料同化 | ▪截斷誤差 | ▪剛體邊界條件 | ▪運動學邊界條件 | ▪海綿邊界條件 |
| ▪初估值 | ▪初始條件 | ▪時間平滑 | ▪空間平滑 | ▪自動資料處理 |
| ▪結構函式 | ▪經驗正交函式 | ▪正交函式 | ▪分離顯式格式 | ▪人造[站]資料 |
| ▪靜力檢查 | ▪差分格式 | ▪顯式差分格式 | ▪向前差分 | ▪交錯格線 |
| ▪交錯格式 | ▪套格線 | ▪張弛法 | ▪假譜方法 | ▪逐步訂正法 |
| ▪最優插值法 | ▪插值法 | ▪統計插值法 | ▪復相關 | ▪復相關係數 |
| ▪人機結合天氣預報 | ▪人機結合 | ▪有限區模式 | ▪有限區細格線模式 | ▪統計動力預報 |
| ▪診斷分析 | ▪大氣強迫 | ▪診斷方程 | ▪診斷模式 | ▪判別分析 |
| ▪決策分析 | ▪決策樹[形圖] | ▪數值模擬 | ▪動力相似 | ▪數值積分 |
| ▪數值實驗 | ▪地圖[放大]因子 | ▪地圖投影 | ▪大氣環流模式 | ▪正壓模式 |
| ▪斜壓模式 | ▪原始方程 | ▪控制方程 | ▪正溫[大氣]模式 | |
昆蟲生態學
| ▪生態系統 | ▪生物因子 | ▪非生物因子 | ▪自養生物 | ▪異養生物 |
| ▪生物地球化學循環 | ▪能量流動 | ▪食物鏈 | ▪食物網 | ▪負二項分布 |
| ▪營養級 | ▪生態錐體 | ▪能量錐體 | ▪生物量錐體 | ▪穩態 |
| ▪生物放大 | ▪環境容量 | ▪環境阻力 | ▪環境適度 | ▪生境 |
| ▪小生境 | ▪生境選擇 | ▪邊緣生境 | ▪生態分布 | ▪斑塊 |
| ▪生態幅[度] | ▪生態閾值 | ▪生態優勢 | ▪生態適應 | ▪生態[種]發生 |
| ▪生態平衡 | ▪生態對策 | ▪生態演替 | ▪生態位 | ▪生態位分化 |
其他科技名詞
| ▪生態位重疊 | ▪生物氣候圖 | ▪昆蟲群落 | ▪群落交錯區 | ▪邊緣效應 |
| ▪種群生態學 | ▪種群 | ▪種群結構 | ▪種群密度 | ▪種群增長 |
| ▪邏輯斯諦增長 | ▪生物潛力 | ▪生物障礙 | ▪活食者 | ▪競爭 |
| ▪食物競爭 | ▪嗜食性 | ▪限制因子 | ▪密度制約因子 | ▪非密度制約因子 |
| ▪時滯 | ▪種群動態 | ▪種群調節 | ▪種群波動 | ▪種群衰退 |
| ▪種群萎縮 | ▪最小存活種群 | ▪種群數量統計 | ▪序貫抽樣法 | ▪雙重抽樣法 |
| ▪隨機抽樣法 | ▪分層隨機抽樣法 | ▪系統抽樣法 | ▪地理分布 | ▪多型群聚 |
| ▪荒漠類群 | ▪沙棲類群 | ▪生態型 | ▪生態表型 | ▪生態梯度 |
| ▪異質種群 | ▪共位群 | ▪異域分布 | ▪生物隔離 | ▪地理隔離 |
| ▪生殖隔離 | ▪生態隔離 | ▪生態效率 | ▪能量收支 | ▪生命表 |
| ▪年齡特徵生命表 | ▪時間特徵生命表 | ▪出生率 | ▪死亡率 | ▪存活率 |
| ▪存活曲線 | ▪關鍵因子分析 | ▪世代時間 | ▪生育力 | ▪繁殖力 |
| ▪光周期 | ▪光周期現象 | ▪淨繁殖率 | ▪周限增長率 | ▪內稟增長力 |
| ▪世代重疊 | ▪多度 | ▪發育起點溫度 | ▪有效積溫法則 | ▪日·度 |
| ▪均勻分布 | ▪隨機分布 | ▪泊松分布 | ▪核心分布 | ▪K 選擇 |
| ▪K 對策昆蟲 | ▪r 選擇 | ▪r 對策昆蟲 | ▪優勢度 | ▪優勢種 |
| ▪地方種 | ▪本地種 | ▪外來種 | ▪關鍵種 | ▪入侵種 |
| ▪廣幅種 | ▪狹幅種 | ▪瀕危種 | ▪稀有種 | ▪捕食作用 |
| ▪捕食者 | ▪獵物 | ▪功能反應 | ▪數值反應 | ▪逃避對策 |
| ▪寄生[現象] | ▪內寄生 | ▪外寄生 | ▪專性寄生 | ▪兼性寄生 |
| ▪擬寄生物 | ▪假寄生 | ▪共寄生 | ▪重寄生 | ▪自復寄生 |
| ▪宿主 | ▪二次寄生 | ▪三次寄生 | ▪群寄生 | ▪獨寄生 |
| ▪單主寄生 | ▪轉主寄生 | ▪群居寄生 | ▪盜食寄生 | ▪宿主專一性 |
| ▪社會性昆蟲 | ▪食物異級 | ▪級 | ▪真社會性昆蟲 | ▪類社會性昆蟲 |
| ▪亞社會性昆蟲 | ▪半社會性昆蟲 | ▪準社會性昆蟲 | ▪雜合群體 | ▪復巢 |
| ▪蟻播 | ▪交哺現象 | ▪蜂糧 | ▪覓食策略 | ▪最優採食 |
| ▪蔽身巢 | ▪共生 | ▪共存 | ▪內共生 | ▪外共生 |
| ▪類共生 | ▪同棲共生 | ▪盜食共生 | ▪互利共生 | ▪偏利共生 |
| ▪偏害共生 | ▪無益共生 | ▪蕈巢共生 | ▪蟻客共生 | ▪守護共生 |
| ▪賓主共棲 | ▪偶然共棲 | ▪客棲 | ▪擬客棲 | ▪客蟲 |
| ▪擬態客蟲 | ▪中性客蟲 | ▪寄食昆蟲 | ▪食客 | ▪蟻客 |
| ▪蟻盜 | ▪蟻菌瘤 | ▪蟲道菌圃 | ▪蟻冢昆蟲 | ▪滯育 |
| ▪專性滯育 | ▪兼性滯育 | ▪過冷卻點 | ▪光期 | ▪暗期 |
| ▪生殖滯育 | ▪低溫滯育 | ▪耐寒性 | ▪擴散 | ▪遷飛 |
| ▪遷出 | ▪遷入 | ▪進化對策 | ▪趨同性 | ▪趨異性 |
| ▪多色現象 | ▪同色現象 | ▪抗生作用 | ▪排拒作用 | ▪異種化感 |
| ▪蟲癭 | ▪間生態 | ▪有害生物綜合治理 | ▪經濟閾值 | ▪生態治理 |
| ▪自然控制 | ▪自然抑制 | ▪雄蟲滅絕 | ▪助增釋放 | ▪保育 |
| ▪就地保育 | ▪異地保育 | ▪檢疫 | ▪檢疫區 | ▪臨界光周期 |
水力學、河流動力學、海岸動力學
| ▪流體力學 | ▪水靜力學 | ▪水動力學 | ▪連續介質 | ▪流體 |
| ▪流體質點 | ▪理想流體 | ▪黏性流體 | ▪牛頓流體 | ▪非牛頓流體 |
| ▪賓厄姆體 | ▪質量力 | ▪[表]面力 | ▪靜水壓力 | ▪動水壓力 |
| ▪黏滯性 | ▪黏滯係數 | ▪表面張力 | ▪毛細管壓力 | ▪流體壓縮性 |
| ▪壓強 | ▪水頭 | ▪水頭線 | ▪位置水頭 | ▪壓強水頭 |
| ▪流速水頭 | ▪慣性水頭 | ▪測壓管 | ▪測壓管水頭 | ▪浮力 |
| ▪浮體 | ▪潛體 | ▪浮心 | ▪定傾中心 | ▪流線 |
| ▪跡線 | ▪過水斷面 | ▪元流 | ▪恆定流 | ▪總流 |
其他科技名詞
| ▪非恆定流 | ▪漸變流 | ▪急變流 | ▪均勻流 | ▪非均勻流 |
| ▪勢流 | ▪渦流 | ▪渦量 | ▪渦線 | ▪渦通量 |
| ▪渦管 | ▪渦絲 | ▪勢渦 | ▪強迫渦 | ▪速度環量 |
| ▪流速勢函式 | ▪流函式 | ▪源 | ▪匯 | ▪等流函式線 |
| ▪等勢線 | ▪流網 | ▪層流 | ▪紊流 | ▪脈動 |
| ▪脈動流速 | ▪壓力脈動 | ▪紊流強度 | ▪時均流動 | ▪猝發 |
| ▪雷諾應力 | ▪紊動黏滯係數 | ▪混合長度 | ▪邊界層 | ▪分離 |
| ▪黏性底層 | ▪水流阻力 | ▪摩擦阻力 | ▪形狀阻力 | ▪阻力係數 |
| ▪升力 | ▪尾跡 | ▪渦街 | ▪摩阻流速 | ▪駐點 |
| ▪水頭損失 | ▪沿程水頭損失 | ▪局部水頭損失 | ▪粗糙係數 | ▪[水力]光滑壁面 |
| ▪水力光滑區 | ▪[水力]粗糙壁面 | ▪水力粗糙區 | ▪阻力平方區 | ▪孔口 |
| ▪堰流 | ▪管流 | ▪管網 | ▪明渠水流 | ▪有壓流 |
| ▪無壓流 | ▪沿程變數流 | ▪管道明滿流 | ▪水擊 | ▪濕周 |
| ▪水力半徑 | ▪稜柱體明槽 | ▪正常水深 | ▪斷面比能 | ▪臨界水深 |
| ▪緩流 | ▪急流 | ▪臨界流 | ▪臨界底坡 | ▪陡坡 |
| ▪緩坡 | ▪水面線 | ▪水力坡降 | ▪水躍 | ▪共軛水深 |
| ▪臨界水躍 | ▪淹沒水躍 | ▪遠驅水躍 | ▪波狀水躍 | ▪水跌 |
| ▪薄壁堰 | ▪寬頂堰 | ▪實用堰 | ▪流量係數 | ▪行近流速 |
| ▪收縮斷面 | ▪收縮係數 | ▪消能 | ▪孔板 | ▪鏇輥 |
| ▪折衝水流 | ▪高速水流 | ▪空化 | ▪空化數 | ▪空蝕 |
| ▪摻氣 | ▪衝擊波 | ▪空泡流 | ▪霧化 | ▪多孔介質 |
| ▪滲流 | ▪滲流阻力 | ▪射流 | ▪羽流 | ▪浮射流 |
| ▪擴散 | ▪分子擴散 | ▪紊動擴散 | ▪離散 | ▪分層流 |
| ▪兩相流 | ▪雷諾數 | ▪弗勞德數 | ▪相似準則 | ▪風洞 |
| ▪減壓箱 | ▪循環水洞 | ▪測針 | ▪畢託管 | ▪普雷斯頓管 |
| ▪文丘里流量計 | ▪彎頭流量計 | ▪電磁流量計 | ▪轉子流量計 | ▪熱絲流速計 |
| ▪熱膜流速儀 | ▪雷射測速儀 | ▪本氏管 | ▪河流泥沙 | ▪侵蝕 |
| ▪輸移 | ▪沉速 | ▪絮凝 | ▪起動流速 | ▪沙波 |
| ▪沙紋 | ▪沙壟 | ▪水流挾沙能力 | ▪輸沙率 | ▪挾沙水流 |
| ▪異重流 | ▪沖刷 | ▪淤積 | ▪輸沙平衡 | ▪河流地貌 |
| ▪河谷 | ▪河漫灘 | ▪主流線 | ▪動力軸線 | ▪河勢 |
| ▪副流 | ▪回流 | ▪河床演變 | ▪河型 | ▪順直型河道 |
| ▪彎曲型河道 | ▪分汊型河道 | ▪遊蕩型河道 | ▪河流過渡段 | ▪河彎 |
| ▪邊灘 | ▪橫比降 | ▪橫向環流 | ▪深槽 | ▪自然裁彎 |
| ▪牛軛湖 | ▪撇彎切灘 | ▪河流節點 | ▪磯頭 | ▪散灘 |
| ▪串溝 | ▪漣子水 | ▪淦 | ▪泡漩 | ▪漿河 |
| ▪揭河底 | ▪造床流量 | ▪河相關係 | ▪普遍沖刷 | ▪局部沖刷 |
| ▪溯源沖刷 | ▪河床質粗化 | ▪回水變動區 | ▪海岸 | ▪海岸帶 |
| ▪海灘 | ▪海岸地貌 | ▪基岩海岸 | ▪淤泥質海岸 | ▪海岸橫向泥沙運動 |
| ▪海岸沿岸輸沙率 | ▪岸灘平衡剖面 | ▪暴風浪型海岸 | ▪涌浪型海岸 | ▪穩定平衡海岸線 |
| ▪河口混合 | ▪含鹽度 | ▪鹽水楔 | ▪攔門沙 | ▪波浪 |
| ▪波峰 | ▪波谷 | ▪波高 | ▪波長 | ▪波周期 |
| ▪波速 | ▪波數 | ▪波陡 | ▪波向 | ▪波向線 |
| ▪波峰線 | ▪波浪中線 | ▪波群 | ▪風成波 | ▪前進波 |
| ▪自由波 | ▪強制波 | ▪深水波 | ▪淺水波 | ▪微幅波 |
| ▪斯托克斯波 | ▪余擺線波 | ▪橢圓餘弦波 | ▪長波 | ▪短波 |
| ▪駐波 | ▪船行波 | ▪孤立波 | ▪規則波 | ▪不規則波 |
| ▪混合浪 | ▪涌浪 | ▪勢波 | ▪波浪破碎 | ▪近破波 |
| ▪遠破波 | ▪波浪繞射 | ▪波浪反射 | ▪波浪折射 | ▪波浪變形 |
| ▪波能 | ▪波能譜 | ▪波壓力 | ▪波浪爬高 | ▪波玫瑰圖 |
| ▪潮汐 | ▪天文潮 | ▪風暴潮 | ▪日潮 | ▪半日潮 |
| ▪混合潮 | ▪大潮 | ▪小潮 | ▪分潮 | ▪漲潮 |
| ▪落潮 | ▪平潮 | ▪潮位 | ▪潮差 | ▪海平面 |
| ▪引潮力 | ▪海嘯 | ▪潮流 | ▪往復潮流 | ▪鏇轉潮流 |
| ▪余流 | ▪憩流 | ▪沿岸流 | ▪漂流 | ▪離岸流 |
| ▪近岸環流 | ▪潮流界 | ▪潮區界 | ▪潮間帶 | |
物理名詞
| 測量 |
|
|---|
| 運動 |
|
|---|
| 熱 |
|
|---|
| 光 |
|
|---|
| 磁 |
|
|---|
| 聲音 |
|
|---|
| 力 |
|
|---|
| 電 |
|---|
擴散
物理學術語
擴散是物質內質點運動的基本方式,當溫度高於絕對零度時,任何物系內的質點都在作熱運動。當物質內有梯度(化學位、濃度、應力梯度等)存在時,由於熱運動而導致質點定向遷移即所謂的擴散。因此,擴散是一種傳質過程,巨觀上表現出物質的定向遷移。在氣體和液體中,物質的傳遞方式除擴散外還可以通過對流等方式進行;在固體中,擴散往往是物質傳遞的唯一方式。擴散的本質是質點的無規則運動。晶體中缺陷的產生與複合就是一種巨觀上無質點定向遷移的無序擴散。晶體結構的主要特徵是其原子或離子的規則排列。然而實際晶體中原子或離子的排列總是或多或少地偏離了嚴格的周期性。在熱起伏的過程中,晶體的某些原子或離子由於振動劇烈而脫離格點進入晶格中的間隙位置或晶體表面,同時在晶體內部留下空位。顯然,這些處於間隙位置上的原子或原格點上留下來的空位並不會永久固定下來,它們將可以從熱漲落的過程中重新獲取能量,在晶體結構中不斷地改變位置而出現由一處向另一處的無規則遷移運動。在日常生活和生產過程中遇到的大氣污染、液體滲漏、氧氣罐泄漏等現象,則是有梯度存在情況下,氣體在氣體介質、液體在固體介質中以及氣體在固體介質中的定向遷移即擴散過程。由此可見,擴散現象是普遍存在的。
晶體中原子或離子的擴散是固態傳質和反應的基礎。無機材料製備和使用中很多重要的物理化學過程,如半導體的摻雜、固溶體的形成、金屬材料的塗搪或與陶瓷和玻璃材料的封接、耐火材料的侵蝕等都與擴散密切相關,受到擴散過程的控制。通過擴散的研究可以對這些過程進行定量或半定量的計算以及理論分析。無機材料的高溫動力學過程——相變、固相反應、燒結等進行的速度與進程亦取決於擴散進行的快慢。並且,無機材料的很多性質,如導電性、導熱性等亦直接取決於微觀帶電粒子或載流子在外場——電場或溫度場作用下的遷移行為。因此,研究擴散現象及擴散動力學規律,不僅可以從理論上了解和分析固體的結構、原子的結合狀態以及固態相變的機理;而且可以對無機材料製備、加工及套用中的許多動力學過程進行有效控制,具有重要的理論及實際意義。
生物學術語
生物個體或其傳布體(如孢子、種子)向其他地域傳布的過程。各種生物都能通過不同的擴散方式擴大棲居範圍,以便獲得更多的資源,並增加其自身種群規模。生物從一地區傳布至另一地區是常見的自然現象,這種異地間的物種交流,促進了各地生物群落的演替,增加了各地區物種的多樣性。大範圍的擴散結合地理隔離是物種分化的重要條件;對物種來說,新的環境意味著不同的自然選擇機制,它將改變種群的基因頻率,因此擴散是促進生物種系進化的重要因素之一。火山爆發毀滅原地生物後,風和洋流會帶來新的生命,各種微生物、植物和動物相繼在此定居。先擴散來的是對異常環境耐受力高的先驅物種,繼之是在安定環境裡競爭能力強的物種,它們在前者開拓的基礎上建立起穩定的群落。
生物擴散包括主動(自力)和被動(借外力)兩類。缺乏行動能力的微生物和植物大多靠被動傳布〔見傳播(植物)〕,動物則多行主動擴散。
主動擴散 生物的生存繁衍需要空間、食物和配偶等條件。生物個體過於稠密時,對空間的需要不易滿足,食源不足及環境惡化會限制種群的發展。與植物相比,動物具有主動尋找生存空間、食源和躲避危險的能力。
動物的主動擴散多發生在生殖期。許多鳥、獸在交配季節四處尋找交配和生育的場所,當其出生地附近的空間多被占據時,就被迫遠距離擴散。其中多數個體往往會因遇到不利環境而死亡,但只要少數個體遇到食源豐富且天敵稀少的合適生境,便可能建立起新的種群,使種的分布區擴大。
許多昆蟲也有很大的擴散能力,而且有些種還表現出多態現象,即同一物種中存在兩種或多種形態和行為均不相同的類型,其中之一專營擴散,具翅或具其他有利於擴散的結構。有的多態現象為遺傳性的,如灰線小卷蛾有兩類不同的遺傳型,一類的幼蟲不耐密集而易擴散,另一類則否;兩者共存互不替代,但數量比例隨種群密度而變。非遺傳性的多態現象則有不同的基礎,有的是發育性的,如蝴蝶、蜻蜓及蜂類的無翅幼蟲取食而不擴散,成蟲則擴散而極少取食。有些昆蟲如蜉蝣的成蟲,甚至完全不取食,僅擴散、交尾、產卵直至死亡。蟻和白蟻的情況也類似。這類社群性昆蟲中,不同社群成員(不同階級)的結構與行為特化,各執行不同的職能,僅有翅型負責擴散到新區域尋找新的繁殖基地。這兩類事例都屬於常規的分工現象,一個是幼蟲與成蟲間的分工,另一個是社群成員間的分工。但飛蝗的多態現象則屬非常規性的,只有當種群密度增加、食物缺乏時,才由散居型經過型變轉化為群居型(即擴散型),造成大規模的擴散,所至之處穀草被其掠食一空。
植物中也有主動擴散的事例,如許多豆科植物果實成熟時果莢裂開將種子散落出去;鳳仙花的果實成熟後,若受外力觸碰,果皮即向內捲縮將種子彈出。不過這種擴散的能力不高,傳播範圍有一定局限。
被動擴散 主要依賴氣流(風)、水流或其他生物的攜帶。
風力傳布 微生物和微小的孢子、種子很容易由風傳送,甚至一些較大的動物也能被大風卷帶很遠。在大洋中部的上空,曾收集到昆蟲和蜘蛛。標記重捕實驗證實,粘蟲可隨3、4月份在中國南方盛行的西南氣流成群遷飛,從華南諸省擴散到江淮流域。還證實,每年6月在日本稻田中褐飛虱數量突增,它們是從中國隨風傳入的。
無翅幼蟲的擴散常以風力傳送為主。蓑蛾幼蟲在母蟲護囊內孵化之後,成群從囊下排泄孔鑽出,吐絲下垂,隨風向四周擴散。有的植物的種子小如粉塵(如蘭科植物),有的植物種子具翅(槭、榆等)、毛(柳、蒲公英等)和氣球狀囊(如酸漿)等特化結構,這都有利於隨風擴散。
水流傳布 借水流擴散的陸地生物必須能浮在水上,不易被水浸透,並能保持內部的滲透平衡。有些專借水流擴散的植物果實表面具不易透水的蠟質或纖維,其組織內含氣,故體輕而不沉。椰子的表面具發達的纖維層,能在水面長期漂流。
巨大的洋流都有固定的行程,因此河口或淡水的水生動物,甚至某些無飛翔能力的陸生動物都可能隨洋流由一個大陸擴散至另一個大陸。有些動物雖會游泳,但其遠距離擴散則可能是隨漂木、浮冰實現的。
動物傳布 寄生生物是借寄主擴散的,有的寄生生物在其生活史的各個階段寄生於不同的寄主。轉換寄主的過程或為主動或為被動。有的寄生物(如血吸蟲)借本身的行動尋找下一個寄主,而有的(如瘧原蟲)則完全被動地擴散,蚊蟲吸食不同寄主個體的血液時瘧原蟲隨之侵染新的寄主。
很多生物依附在其他生物的體表擴散。依附的方式很多:有些植物的果實和種子具刺或鉤(如蒼耳的果實),還有的具粘性分泌物(如亞麻種子);一些小昆蟲,蟎類等常粘附在脊椎動物的皮毛上,也有些小動物粘附在候鳥的足上。
有的植物種子經動物取食而擴散。例如草食動物經常吃下大量種子,這些種子可隨糞便排至多處。很多肉質果實憑其色、香、味吸引動物取食,其種子或被吐出或隨糞便排出。這些種子一般均能抵抗動物消化液的侵蝕,有些種子只有在種皮經消化道作用後才能萌發。
人類也是重要的生物擴散因子。長途的人貨流通有意無意地擴散了大量生物。棉花的主要害蟲棉紅鈴蟲原產印度,20世紀30年代傳入中國沿海地區,現在除新疆外已遍及中國各棉區。此外,人類還有意識地引種或培育很多優良的穀物、林木和家禽家畜,並在全世界範圍推廣。
擴散力的進化 在自然界,擴散力存在兩種截然不同的進化趨勢,一種是擴散力增強,另一種是減弱甚或消失。例如加拿大不列顛哥倫比亞省省會維多利亞附近的溫哥華島的西部天幕毛蟲,在其種群中,擴散力弱的個體在生境附近產卵,後代常死於惡劣氣候;擴散力強的個體向遠處產卵,生於較有利環境中的後代可能存活,並且大多是活潑個體。如此代代選擇,該種的擴散能力不斷增強。另一方面,如果靠擴散找到未開拓的適宜環境的機會很小時,自然選擇將對抗擴散。例如在海島和山巔環境中不會飛的鳥和昆蟲種數比平坦的大陸上要多。達爾文在考察馬德拉群島時注意到,島上550種甲蟲中有200種有翅卻缺乏飛翔能力,而當地29個屬甲蟲中有23個屬於狂風時潛藏,天氣較好時才外出活動。達爾文認為,這種情況主要是因為善飛個體在飛行中可能被狂風吹入大海,而翅發育不全或習性怠惰的個體,反能更多地存留下來。動物地理學家P.J.達林頓認為,遇敵時翅膀有助於逃生,但海島上天敵很少或沒有,翅就成為無用之物。與此類似,島上植物的擴散力也明顯下降或幾乎完全消失。有些植物果實變大,但用於傳布的附器(如倒鉤)卻未相應增加,有的種子附器甚至退化,這類種子或果實的數量也常相應地減少。這種進化變異可減少向島外的無效擴散,而個體增大則可更多地儲存營養,反而有利於幼苗生長。
人工選擇也可改變擴散能力。例如人類栽培農作物要求的是高收穫量,而不是高擴散力。目前種植的小麥、水稻等穀物的種子散落性已消失,亞麻和罌粟蒴果的開裂機制也已退化。
擴散現象的數學模型 根據觀察種子散落的情況和用標記重捕法進行的昆蟲擴散試驗,人們發現,隨著擴散距離的增加,可能找到的傳布體的數量按指數比率下降。可用經驗式表示為:
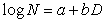
種子散落屬於被動擴散;有些小昆蟲飛翔力不強,方向和距離也基本是隨機的,所以也表現上述規律。
擴散是研究種群數量變動的重要因子。在生產實踐中,害蟲和雜草的擴散會給人類健康和農牧業帶來損害。因此,了解生物擴散的特性,對防治有害生物有重要意義。許多細菌性和病毒性人畜疾病及作物病害的防治,往往取決於切斷病原體的傳布途徑。稻縱卷葉螟和褐飛虱是中國南方成群長距離擴散的重要水稻害蟲,田間種群數量經常突增或突減,掌握其擴散規律便可預測它的發生量和發生期。利用捕食性天敵防治害蟲時,必須研究天敵和害蟲在各種環境(包括不同的地形、植物長勢、氣候變化等)中的擴散能力、傳布方向和可能的傳布距離等,以便確定人工釋放天敵的數量、野外釋放點布局和釋放點高度等。
配圖
相關連線
| ▪電荷 | ▪正極 | ▪電壓 | ▪帶電體 | ▪負極 |
| ▪伏特 | ▪摩擦起電 | ▪電路 | ▪伏特計 | ▪正電荷 |
| ▪通路 | ▪電阻 | ▪負電荷 | ▪開路 | ▪歐姆 |
| ▪導體 | ▪短路 | ▪變阻器 | ▪絕緣體 | ▪電路圖 |
| ▪滑動變阻器 | ▪半導體 | ▪串聯 | ▪歐姆定律 | ▪電源 |
| ▪並聯 | ▪電功 | ▪導線 | ▪電量 | ▪電功率 |
| ▪電鍵 | ▪電流 | ▪額定電壓 | ▪乾電池 | ▪安培 |
| ▪額定電功率 | ▪蓄電池 | ▪安培計 |
