
簡介
埃及棘龍(屬名:Spinosaurus)意思為“有棘的蜥蜴”,是種巨型的獸腳亞目恐龍,生存於白堊紀中期到晚期的非洲,約為11200萬年到6500萬年前。棘龍最初是在1912年由德國古生物學家恩斯特·斯特莫(Ernst Stromer)發現於埃及,並在1915年加以敘述、命名。最初的標本和最大標本在第二次世界大戰期間被盟軍的飛機轟炸所摧毀,但最近幾年相繼發現了一些不完整頭顱骨、脖子骨頭、神經弓、牙齒、爪子、腳指頭、胳膊骨頭。如今仍不確定棘龍屬有一個種還是兩個種,其中最著名的種是發現於埃及的埃及棘龍(S. aegyptiacus),而第二個種是摩洛哥棘龍(S. marocannus),化石發現於摩洛哥。更有很多獨立的牙齒、頜骨碎塊、爪子等部分骨骼化石被發發現。還有一些保存很良好的化石落入私人收藏家手裡
。
棘龍的背部有明顯的長棘,是由脊椎骨的神經棘延長而成,長度可達6米,長棘之間可能有皮膚連結,形成一個帆狀物;然而有些科學家認為這些長棘是由肌肉復蓋著,形成隆肉或是背脊。對於這帆狀物的功能,如今已有數種看法,包括調節體溫、視覺展示物。棘龍的頭顱骨長而低矮,外形類似上龍類,棘龍被認為是以小到大型鳥臀目恐龍和小中型蜥腳類為食的頂級掠食者,但偶爾也食魚類。一項針對棘龍科牙齒的氧同位素組成研究,推測棘龍是半水生動物,可在陸地、水域中生存,類似現代棕熊。根據最近的估計,棘龍是已知最大型的肉食性恐龍和獸腳亞目恐龍,也是已知歷史上最大的陸地食肉動物,甚至大於霸王龍和蠻龍。這些推算顯示棘龍身長12-19米,臀高2.7-4米,高度約為5-6米,體重為4-18噸,但也有少數研究人員不接受這些數據。根據研究,棘龍的平均咬合力為2噸,掌擊力量可達到2噸。
別名:棘龍、脊背龍(Spinosaurus並無標準譯名,兩個譯名皆可,而“棘龍”使用更廣)
生存時期:11200萬年前(中生代早白堊紀)-6500萬年前(中生代晚白堊紀)
科學分類:
界:動物界Animalia
門:脊索動物門Chordata
綱:蜥形綱Sauropsida
總目:恐龍總目Dinosauria
目:蜥臀目Saurischia
亞目:獸腳亞目Theropoda
總科:斑龍超科Megalosauroidea
科:棘龍科Spinosauridae
亞科:棘龍亞科Spinosaurinae
屬:棘龍屬Spinosaurus
體型數據:
體全長:12-17米
最長:18米
臀高 2.7-4米
胳膊長:2米
爪全長:40厘米
帆高:2米
頭長:2米
體重:4-15噸
時代:白堊紀中晚期
年代距今約(living age):11200萬年前-6500萬年前
生存地區:非洲北部的突尼西亞、摩洛哥、阿爾及利亞、埃及、利比亞,可能還有尼日和肯亞
分類
 埃及棘龍
埃及棘龍棘龍科的名稱來自於棘龍屬,棘龍科可分成重爪龍亞科、棘龍亞科。重爪龍亞科包含:歐洲的重爪龍、尼日的
似鱷龍和脊飾龍以及寮國的魚獵龍,還有隻有很碎化石的鱷龍(英國)。棘龍亞科還包括:非洲的棘龍、巴西的激龍與奧沙拉龍、泰國的暹羅龍和中國的中國上龍(後兩者如今只有發現很破碎的化石);此外,還有基礎棘龍科東非龍。棘龍與奧沙拉龍的親緣關係最接近,兩者都屬於棘龍亞科,都擁有筆直、無鋸齒邊緣的牙齒,牙齒間距寬,棘龍的每邊上頜骨有12顆牙齒;重爪龍亞科有彎曲、鋸齒邊緣的牙齒,牙齒數量較多,每邊上頜骨約有30顆牙齒。此外,棘龍科的全體成員都擁有尖利有力的大型指爪,長度從20-60厘米不等。
在2003年,奧利佛·勞赫(Oliver Rauhut)提出斯特莫所發現的棘龍正模標本是個嵌合體,背椎是來自於類似高棘龍的鯊齒龍科恐龍,而齒骨來自於類似重爪龍的大型獸腳類恐龍。然而,這個論點已被最近的研究所否認。
發現與種
棘龍屬目前有兩個已命名種,分別為埃及棘龍(Spinosaurus aegyptiacus)、摩洛哥棘龍(S. marocannus)。
棘龍的第一個化石是在1912年發現於埃及西部的拜哈里耶綠洲,並由德國古生物學家恩斯特·斯特莫(Ernst Stromer)在1915年命名為模式種埃及棘龍。之後在拜哈里耶綠洲也發現了其他的化石碎片,包含脊椎與後肢,恩斯特·斯特莫在1934年將這些化石標名為"SpinosaurusB"。斯特莫認為這些新發現化石有足夠差異,因而歸類於另外一個種;而如今其他研究人員證實這些差異,這些新發現化石可能與
鯊齒龍有關,或是與斯基瑪薩龍有關聯。有些棘龍化石在運送到德國慕尼黑德意志博物館的運送過程中遭到損毀,而這些骨頭在1944年的盟軍轟炸中遭到破壞、遺失。
摩洛哥棘龍是在1996年由戴爾·羅素(Dale Russell)所研究、命名,羅素是根據頸椎長度而將它們建立為一個新種。羅素比較頸椎椎體的長度、椎體後緣的高度,發現埃及棘龍的比例為1.1,而摩洛哥棘龍的比例為1.5。然而,數名研究人員認為摩洛哥棘龍的頸椎長度只是個體間的變化,而埃及棘龍的正模標本已被轟炸破壞,因此無法比對兩者是否為相同動物,而且無法確定摩洛哥棘龍的頸椎是第幾節,而其他科學家仍主張它們是另一有效種。如今大部分古生物學家認為摩洛哥棘龍是個疑名,或者是埃及棘龍的次異名。
標本
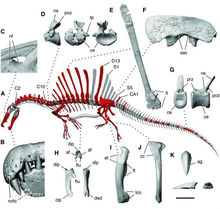
已有數個棘龍的編號標本被敘述,另外還有更多無標號的化石,在科學研究機構和私人收藏手裡。其中第一個標本和最大標本在第二次世界大戰中遭到摧毀,但是已留下了詳細的敘述與繪畫。藉由測量其他棘龍科恐龍的體型,可以估算出這些棘龍個體的體型大小。以下的估算數據來自於獸腳亞目資料庫以及克里斯提亞諾·達魯·沙索(Cristiano Dal Sasso)等人在2005年的研究。
BSP1912VIII19標本
 埃及棘龍
埃及棘龍由恩斯特·斯特莫在1915年所敘述,化石發現於埃及的拜哈里耶組,是埃及棘龍的正模標本,原本的編號是IPHG 1912 VIII 19。該標本由一個上頜骨碎片、一個不完整齒骨(75厘米長)、19顆牙齒(6.2厘米長)、2個不完整頸椎、7個背椎(19到20厘米)、肋骨、以及1個尾椎。該標本的下頜有1.34米長。最長的脊椎神經棘有1.69米,長莫認為這個化石發現於早森諾曼階,約9700萬年前。估測14米長重8.6噸。這個標本存放於慕尼黑古生物博物館,之後在第二次世界大戰中遭到盟軍轟炸摧毀,僅存一個腳趾頭化石。恩斯特·斯特莫當年曾做出詳細的標本素描圖,他的孫子在1995年將素描圖捐給慕尼黑古生物博物館。Joshua B. Smith、Matthew C. Lamanna根據兩張重新發現的標本照片,發現斯特莫的標本素描圖有少許不正確。在2003年,奧利佛·勞赫提出這個標本是個嵌合體,齒骨來自於重爪龍或似鱷龍,脊椎與神經棘來自於類似高棘龍的鯊齒龍科,但是達魯·沙索和絕大部分科學家則堅決反對這個毫無根據的理論。
BM231標本
由埃瑞克·比弗托(Eric Buffetaut)與M. Ouaja在2002年所敘述,該標本發現於突尼西亞的雪尼尼組,地質年代為早阿爾比階,約11200萬年前,是棘龍屬年代最早的標本。化石由一個齒骨前部構成,可發現四個齒槽、兩個部分牙齒。外形非常類似埃及棘龍的失蹤標本。該化石標本在棘龍屬里出奇的小而且年代比其他標本都早,因此在2010年某些科學家主張建立單獨的突尼西亞棘龍種。一些科學家在2010年主張將這個標本單獨建立突尼西亞棘龍種的申請,已被科學家否認,因為“突尼西亞棘龍”的模式標本(也是唯一標本),無任何可鑑定的獨特特徵,無法將其與其他兩個種(埃及棘龍和摩洛哥棘龍)區分開來,而且很可能就是埃及棘龍早期種群.體型估測11.5米5.4噸。
UCPC-2標本
同樣由沙索等人在2005年所敘述,發現於摩洛哥的卡瑪卡瑪地層,年代為9700萬年的早森諾曼階。該標本是一個有凹槽的冠飾,由兩塊鼻骨所構成,長度為18厘米
NMC41852
1996年發現在摩洛哥,一個75厘米長的肱骨,但也可能屬一隻超大的似鱷龍。棘龍擁有延長的嘴部、圓椎狀牙齒、以及較高的鼻孔,類似上龍類,顯示它們可能是廣生性物種,以大到中型蜥腳類恐龍、小到大型鳥臀目恐龍為主食。棘龍很可能是種無特定目標、多種食物來源的食肉動物。
MSNMv4047標本
 埃及棘龍
埃及棘龍由米蘭市民自然歷史博物館的克里斯提亞諾·達魯·沙索與他的同事在2005年所敘述,發現於摩洛哥的卡瑪卡瑪地層,年代為9700萬年的早森諾曼階。該標本由前上頜骨、部份上頜骨、以及部份鼻骨所構成,這些部位總長度為98.8厘米,復原頭骨全長為175厘米。
食性
如今的絕大多數說法都認為棘龍主要是陸地頂級掠食者,而不是過去所認為的純粹魚食性動物,以小中型蜥腳類恐龍、小到大型鳥臀目恐龍為主食,也會吃腐肉和魚類甚至翼龍類。棘龍食性的唯一直接證據來自於它們的近親,居住於歐洲與南美洲的重爪龍。曾在重爪龍的胸腔中,發現了一些鱗齒魚(Lepidotes)的鱗片;另一個標本的胃部曾發現幼年禽龍的骨頭。另外,在南美洲發現的一個翼龍類化石上,嵌入者一個棘龍類的牙齒,顯示棘龍類偶爾以這些飛行主龍類為食。棘龍很可能是種無特定目標、多種食物來源的頂級掠食動物,可用白堊紀的大型灰熊來形容它們,平時捕食鳥臀目恐龍和小中型蜥腳類,也會吃別的恐龍剩下的腐肉或者去捕食魚類,甚至會以翼龍類為食。
在2009年,達魯·沙索等人對編號MSNM V4047標本進行X射線計算機斷層成像,發現棘龍的上頜下頜有疑似感應器官的小孔。達魯·沙索等人推測,棘龍可能偶爾會將身體浸泡在水中,頭部半露出水面,並感應周圍的魚類或水邊恐龍的動靜,而進行獵食。
在2010年,一項針對棘龍科牙齒的氧同位素研究,顯示棘龍是半水生動物。這項研究分析棘龍、重爪龍、激龍、暹羅龍的氧同位素組成,並與同時代的其他獸腳類恐龍(例如鯊齒龍)、烏龜、鱷魚互相比較。研究人員發現棘龍牙齒的氧同位素組成,較接近同一地區發現的烏龜、鱷魚,而不類似同一地區的獸腳類牙齒。
棘龍亞科
棘龍屬於棘龍亞科,棘龍亞科是由塞里諾在1998年所建立的。在2004年,並由湯瑪斯·荷茲(Thomas R. Holtz Jr.)等人定義為親緣關係較接近埃及棘龍、而離沃克氏重爪龍較遠的所有物種。該亞科包含:棘龍、奧沙拉龍、激龍、中國上龍、暹羅龍(但後兩個屬存在著一些爭議,因為化石很碎)。該亞科最早化石是1.3億年前的,最晚化石記錄是8400萬年前,分布範圍包括亞洲、非洲、南美洲。1998年最初建立時還包含崇高龍,但2012年研究發現崇高龍和激龍其實是同種動物,於是崇高龍屬被激龍屬吞併。在棘龍科和荊棘龍亞科里,和棘龍關係最近的就是奧沙拉龍了,其次是激龍。棘龍和激龍、奧沙拉龍很接近,而離中國上龍、暹羅龍關係偏遠一點。中國上龍在2008年研究被重新歸入棘龍科棘龍亞科,暹羅龍在2012年重新研究後歸入棘龍亞科。1998年建立了激龍族,包含了激龍和崇高龍,崇高龍屬撤消後該族依然存在,僅包含激龍。和重爪龍亞科相比,棘龍科骨骼更粗壯結實,背帆很大而且突出,食性更廣,頭骨(特別是頜骨)更粗壯和強壯。
牙齒
棘龍的牙齒很長,一般牙齒外露長度在7-8厘米之間,目前發現最大牙齒可達12.5厘米,平均個體的棘龍的咬合力也很強,有3噸以上。
體型
自從棘龍被發現之後,它們成為最長、最大型獸腳類恐龍的候選者之一,但一般大眾對於棘龍的巨大體型並沒有概念,直到棘龍出現在2001年的電影《侏羅紀公園 III》(Jurassic Park III),以及2005年對於它們新標本的研究。弗雷德里克·馮·休尼博士(Friedrich 他們認為棘龍身長為15米,重量為6公噸以上。身長一樣被敘述為15米,但估計的體重數據較低,約4公噸
。
在2005年,克里斯提亞諾·達魯·沙索(Cristiano Dal Sasso)認為棘龍、似鱷龍有類似的身體與頭顱骨比例,而似鱷龍的身長估計約為11米,體重約4.6噸。達魯·沙索等人根據這個數據,而將棘龍身長估計為16到19米,體重為7到9公噸(骨長法)或16~22公噸(3D建模法)。,棘龍科的不同屬是否有不同的頭顱骨形狀,而這將導致不同的身長估計值。
在一個最近的研究中,根據頭顱骨長度來推算比例,發現之前所估計的數據中,身長太長,而體重太重。他們根據頭顱骨的長度為1.5到1.75米,而估計完整為身長12.6到14.3米,而體重為12到20公噸。根據這個數值,棘龍的身長將比鯊齒龍、馬普龍、蠻龍還短,體重也比較輕。但他們的研究遭到批評,原因是他們所選擇的大型獸腳類對照組,此外,他們所重建的棘龍科頭顱骨過短,可能影響數據。需要一些更加完整的化石才能徹底解決這些關於體型上的爭議,尤其是大腿、小腿、骨盆的骨骼化石.土,矮型棘龍取代了高型棘龍。
 埃及棘龍
埃及棘龍種種跡象表明:棘龍是兩棲動物,主要以魚,鱷魚為食。但在旱季時,棘龍也會上岸捕食豪勇龍,皺褶龍,還會和鯊齒龍發生衝突。
2014年9月發現成年棘龍化石。
帆狀物
棘龍擁有獨特的帆狀物,雖然同一時期的該地區其他恐龍也可能發展出類似的背椎結構物,例如鳥腳下目的豪勇龍、蜥腳下目的雷巴齊斯龍,以及生存於南美洲的蜥腳類阿瑪加龍。棘龍的帆狀物也類似異齒龍的背帆;異齒龍是種似哺乳爬行動物,生存於恐龍時代以前的二疊紀,兩者的帆狀物並非同源演化的特徵,早在1915年,恩斯特·斯特莫就認為這些背部結構物支撐者脂肪構成的隆肉。在1997年,Jack Bowman Bailey如今提出棘龍、豪勇龍的背棘並非細棒,而是前後軸寬廣,不同於盤龍目的扁平背帆,而是類似巨角犀、水牛的背脊,。
棘龍的帆狀物功能仍未確定;科學家們已提出數個假設,包含調節體溫、吸引異性的展示物。
這顯示棘龍至少是某種程度的溫血動物,之後有其他研究人員提出反對意見,認為如果棘龍有背帆,它們吸收熱量的效率,將高於散發熱量的效率。他們認為這些有長神經棘的恐龍,背部具有脂肪構成的隆肉,以儲存能量。
許多現代動物的複雜身體結構,在求偶季節時具有吸引異性功能。這些恐龍的帆狀物有相當可能性具有吸引求偶功能,類似孔雀的尾巴。斯特莫假設這些恐龍的雄性與雌性擁有不同大小的神經棘。如果屬實,這些帆狀物可能擁有耀眼的顏色,但這是完全建立於推測上的。
姿勢
在傳統看法裡,棘龍被敘述成二足動物,80年代早期之後,它們被認為至少有時可用四足方式行走。這個論點因為發現了重爪龍而得到支持,重爪龍是棘龍的近親,擁有結實的手臂。在1997年,Jack Bowman Bailey贊成棘龍為可能四足姿勢,這可見於他研究中的新重建圖。根據一個侏羅紀早期的獸腳類足跡化石,它們有可能以手掌側邊接觸地面而休息。
前肢功能
與人類相比,棘龍的肩膀的轉動範圍小的多。它們的手臂無法做出360°的鏇轉幅度,但可後擺至離垂直面約125°,所以肱骨可以後擺至斜微上方。手臂可以前轉至超過垂直面約40°。手臂無法往垂直地往下擺,但可外展至超於水平面15°。與人類相比,手肘的活動範圍也很小,大約只有70°的轉動幅度。手臂無法完全地伸直,也無法大幅彎曲,而肱骨不能做出100°彎曲。橈骨與尺骨互相固定,所以無法如人類的前臂,做出往內側或外側鏇轉的動作。
移動速度
儘管棘龍的後肢只有腳指頭化石被發現,但科學家們仍然根據其近親的腿部推斷並復原了棘龍的腿部骨骼和其腿部的結構。
關於棘龍是否能夠高速奔跑,以及能以多快的速度移動,則有許多互相衝突的研究。針對棘龍的行走極速,科學家們和學者們已提出各種不同的估計值。
帆狀物的功能
棘龍擁有獨特的帆狀物,這是在現代動物身上不復存在的。雖然同一時期的該地區其他恐龍,例如鳥腳下目的豪勇龍、蜥腳下目的雷巴齊斯龍,也可能發展出類似的背椎結構物。棘龍的帆狀物類似二疊紀的似哺乳爬行動物異齒龍的帆狀物,異齒龍生存於恐龍時代以前,兩者的帆狀物並非同源,而是平行演化的結果。棘龍的背部結構物可能較類似駝峰,而非帆狀物;Jack Bowman Bailey如今提出,棘龍的背棘並非細棒,而是前後軸寬廣,類似水牛的背脊,所以棘龍的背棘應是支撐者較厚、較肉質的結構物,而非細的帆狀物,但根據英國科學家的最新推論,白堊紀中期到白堊紀末期時的撒哈拉沙漠很有可能是一片雨林,這就意味著棘龍的帆狀物的作用不太可能像駱駝的駝峰那樣。
棘龍的帆狀物功能仍未確定;科學家們已提出數個假設,包含調節體溫、展示物、捕魚。此外,如此明顯的背部特徵可使棘龍的外表看起來比較大,可威脅其他動物。主流學說仍然是調節體溫。
如果這些帆狀物擁有大量的血管,棘龍可使用帆狀物的大型表面積來吸收熱量。這顯示棘龍至少是部分溫血動物,並生存於夜間氣溫低、冷,但天空通常很少雲的地區。棘龍與豪勇龍被認為為生存於早期撒哈拉沙漠的邊緣,這可解釋上述的功能。這些帆狀物也有可能用來釋放多餘的體溫,而非收集熱量。根據霍爾丹法則(Haldane's principle),與身體體積相比,大型動物反而擁有較小比例的表面積,它們釋放的熱量溫度較高,而吸收的熱量溫度較低。這些恐龍的帆狀物增加相當程度的身體皮膚面積,而且所增加體積最少。此外,如果將這些帆狀物遠離太陽方向,或是以90度迎接較冷的風,這些身處白堊紀非洲的動物將能更有效地降低體溫。
許多現代動物的複雜身體結構,在求偶季節時具有吸引異性功能。這些恐龍的帆狀物有相當可能性具有吸引求偶功能,類似孔雀的尾巴。斯特莫假設這些恐龍的雄性與雌性擁有不同大小的神經棘。 如果屬實,這些帆狀物可能擁有耀眼的顏色,但這是完全建立於推測上的。
最特殊的方式:捕魚
已經有多位科學家提出:棘龍是半水生動物,而且棘龍的帆狀物因為薄而窄,所以極有可能用來捕魚,怎么使用呢?現代動物:很多現代動物都會在捕魚時製造陰影,而魚有一個特性,那就是易被陰影吸引,而這些動物就可以趁機捕食,而且,它們製造陰影物都不重,所以,科學家猜測,這棘背極有可能還會用來捕食魚類
。
這些帆狀物很可能擁有綜合以上功能,平常充當體溫調節器、儲存脂肪能量、在求偶季節時吸引異性、散發體溫、捕魚以及在遭受威脅時充當警告物使用。
分布範圍
棘龍的棲息環境涵蓋北非絕大部分,可能還有西非尼日和東非肯亞但只有部分了解。棘龍生存於當時埃及地區的海岸與潮坪環境,與類似的大型掠食者如 巴哈利亞龍、 鯊齒龍、三角洲奔龍、索倫龍,大型的 泰坦巨龍類 潮汐龍的幼崽,較小型的泰坦巨龍類 埃及龍,9公尺長的鱷類肌鱷,以及 腔棘魚類的Mawsonia。
