
簡介
 反轉錄酶
反轉錄酶這是一種莫洛尼鼠白血病病毒(Moloneymurineleukeminvirus)反轉錄酶(簡稱M-MLVRTase),
這個酶已經作了遺傳性上的改變,即除去了與它聯在一起的核糖核酸酶H活性(一種專門切割DNA與RNA雜交鏈上RNA分子的酶)。這個酶套用於cDNA的第一鏈合成和引物的延伸。這種酶需要鎂離子或錳離子作為輔助因子,當以mRNA為模板時,先合成單鏈DNA(ssDNA),再在反轉錄酶和DNA聚合酶Ⅰ作用下,以單鏈DNA為模板合成“髮夾”型的雙鏈DNA(dsDNA),再由核酸酶S1切成二條單鏈的雙鏈DNA。因此,反轉錄酶可用來把任何基因的mRNA反轉錄成cDNA拷貝,然後可大量擴增插入載體後的cDNA。也可用來標記cDNA作為放射性的分子探針。
逆轉錄酶(M-MLV)從MoloneyMurineLeukemiaVirus分離出來,可用於合成第一鏈cDNA、製作cDNA探針、RNA轉錄、測序和RNA的逆轉錄反應。本酶是通過點突變使RNaseH活性缺失,所以它具有的DNA聚合酶的活性與野生型相同,同時其延伸能力也有顯著提高。逆轉錄酶(M-MLV)一個活性單位定義為在37℃,10min條件下,使1nmol的脫氧核糖核酸摻入酸性沉澱物質所需的酶量。
合成步驟
 反轉錄酶
反轉錄酶1、使用前每個組份輕輕混勻,然後2000rpm離心20s;
2、取滅過菌且無核酸酶的0.2ml離心管,依次加入2~5μgRNAnμL
3、65℃保溫5min,然後冰浴5min;
4、往3步驟中的0.2ml離心管依次加入下列組份
RNase抑制劑(40u/μL)0.5μL
10×M-MLVReactionBuffer2μL
DTT(200mM)1μL
逆轉錄酶(M-MLV)1μL
5、輕輕混勻後,然後2000rpm離心20s;
6、先在37℃保溫1hr,然後70℃保溫15min;
7、上述產物可立即進行下一步的PCR反應或-20℃保存。
主要特點
多反轉錄酶都具有多種酶活性,主要包括以下幾種活性
。
①DNA聚合酶活性;以RNA為模板,催化dNTP聚合成DNA的過程。此酶需要RNA為引物,多為色氨酸的tRNA,在引物tRNA3′-末端以5′→3′方向合成DNA。反轉錄酶中不具有3′→5′外切酶活性,因此沒有校正功能,所以由反轉錄酶催化合成的DNA出錯率比較高
。
②RNase H活性;由反轉錄酶催化合成的cDNA與模板RNA形成的雜交分子,將由RNase H從RNA5′端水解掉RNA分子
。
③DNA指導的DNA聚合酶活性;以反轉錄合成的第一條DNA單鏈為模板,以dNTP為底物,再合成第二條DNA分子
。
除此之外,有些反轉錄酶還有DNA內切酶活性,這可能與病毒基因整合到宿主細胞染色體DNA中有關。反轉錄酶的發現對於遺傳工程技術起了很大的推動作用,目它已成為一種重要的工具酶。用組織細胞提取mRNA並以它為模板,在反轉錄酶的作用下,合成出互補的DNA(cDNA),由此可構建出cDNA文庫(cDNa library),從中篩選特異的目的基因,這是在基因工程技術中最常用的獲得目的基因的方法,詳見基因工程一章
。
質量控制
 反轉錄酶
反轉錄酶1.切口酶:在與200單位酶37oC保溫60分鐘後,超螺鏇質粒保留在90%以上。 2.DNase測定:200單位酶與50ng3H-DNA37oC保溫60分鐘,分解出的放射性低
於1%。
3.RNase測定:200單位酶與50ng3H-RNA37oC保溫60分鐘,分解出的放射性低
於3%。
4.第一鏈cDNA的反應:在標準化的反應中,200單位酶催化從1mg1.2kbRNA或
1mg7.5kbRNA將同位素摻入到cDNA中,通過放射自顯影,對於1.2kb的RNA,產物
cDNA是單一的、全長的條帶。對於7.5kb的RNA產物有部分是全長的條帶。用這二種
RNA放射性轉換到cDNA中有12%。
5.RNaseH活性:200單位酶與polyA:polydT37oC保溫60分鐘,釋放出的放射
性要低於1%。
6.物理純度:在SDS-PAGE上>90%純。
儲存緩衝液:20mMTris-HClpH7.5;200mMNaCl;0.1mMEDTA;1mMDTT;0.01%NP-40(一種
去垢劑)和50%甘油。
反應緩衝液:250mMTris-HClpH8.3;375mMKCl;15mMMgCl2;50mMDTT(以Part#M531A供應)。
使用說明
 反轉錄酶
反轉錄酶M-MLV反轉錄酶比AMV反轉錄酶缺乏連續性,因此要獲得像AMV反轉錄酶反應中產生同樣量的cDNA就要求使用較多單位的M-MLV反轉錄酶。用1微克的mRNA起始合成第一條鏈的cDNA,要用8單位的M-MLV反轉錄酶才相當於1單位的AMV反轉錄酶的作用。
該酶很容易被亞精胺(Spermidine)所抑制,該酶絕對不能用Promega的RiboprobeAMV反轉錄酶5X反應緩衝液,也不能用Promega的RiboclonecDNA第一條鏈緩衝液,因為這二種緩衝液均含有亞精胺。
注意事項
 反轉錄酶
反轉錄酶在進行RT反應之前,應考慮以下幾個方面:
1、RNA
成功的cDNA合成來自高質量的RNA,高質量的RNA至少應保證全長並且不含逆轉錄酶的抑制劑,如EDTA或SDS。在提取RNA的過程中,要特別防止RNase的污染,同時在逆轉錄反應中經常加入RNase抑制劑以增加cDNA合成的長度和產量。RNase抑制劑要在第一鏈cDNA合成反應中,在緩衝液和還原劑(如DTT)存在的條件下加入,因為cDNA合成前的過程會使抑制劑變性,從而釋放結合的可以降解RNA的RNase。蛋白RNase抑制劑僅防止RNaseA,B,C對RNA的降解,並不能防止皮膚上的RNase,因此儘管使用了這些抑制劑,也要小心不要從手指上引入RNase,實驗過程中經常更換新手套。
2、引物的選擇
OligodT
選擇OligodT時,要求RNA必須有PolyA,所以真核生物的mRNA都適用。適合長鏈甚至全長mRNA的RT,所以對RNA樣品的質量要求較高,最好不要有明顯的DNA污染、RNA降解和RNA斷裂。假如想探索新的mRNA進行RT反應,建議推薦使用OligodT引物。使用OligodT引物要比隨機引物和特異性引物的穩定性要好。
隨機引物
適合各種RNA的RT,尤其適合模板豐度很低的情況(比如某個gene表達量很低)。選擇隨機引物時,第一鏈cDNA合成反應中就是以所有的RNA為模板,然後進行PCR反應時設計引物進行特異性擴增。同時要注意隨機引物的量和總RNA量之間的關係,一般建議每5μg總RNA的隨機引物的用量為50ng,如果每5μg總RNA的隨機引物的用量超過250ng,可能會導致小片段產物(<500bp)的增加和長片斷、全長產物產物的降低。
特異性引物
特異性引物只能用你設計引物時的下游引物做RT,引物設計質量影響RT的結果,而且不同引物退火溫度本來就不相同,所以按照說明書按照一個溫度做不是最佳選擇,一般不推薦。
醫學發展
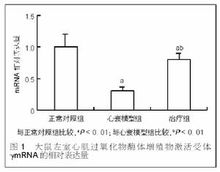
細胞的衰老和老化被認為和染色體末端由重複的DNA(TTAGGG)序列所組成的端粒序列的丟失相關。隨著細胞的每次分裂,端粒會丟失50~200bp,當端粒縮短到一定程度就不再保護染色體免受重組或降解,細胞分裂的控制點就此得到信號而產生作用,可使細胞分裂停止並進入老化過程致細胞死亡。端粒長度的維持即重複序列向染色體末端的添加由端粒酶所催化。端粒酶組成包括RNA組分、端粒酶相關蛋白和端粒酶反轉錄酶。其中端粒酶反轉錄酶對端粒酶活性起關鍵作用。在人體,端粒酶活性存在於幹細胞、生殖細胞、部分有再生能力的體細胞及絕大多數惡性腫瘤組織中。它的高活性是腫瘤細胞惡性增殖的一項重要條件,但其適量表達又具有延長細胞壽命的作用。隨著幹細胞分化的進行,端粒酶活性也隨之降低,至終末分化細胞已無法測到端粒酶。適度上調和維持幹細胞的端粒酶活性,對提高幹細胞體外複製和擴增能力及揭示幹細胞衰老機制都將具有重要意義。近期研究發現,在富含表皮幹細胞的表皮基底層和生長期毛囊中均有端粒酶活性表達,但目前對不同發育階段人表皮幹細胞端粒酶表達特徵仍缺乏相應的了解。弄清楚它們在表皮幹細胞中的表達變化及其規律,將會增加對表皮幹細胞的認識,獲得更多有關表皮幹細胞複製調控的新信息。
成功將人表皮幹細胞體外分離培養的基礎上,對比觀察不同發育階段人表皮幹細胞端粒酶反轉錄酶表達的差異特徵,結果表明人胚胎、少兒、成人皮膚來源的表皮幹細胞均有端粒酶反轉錄酶表達,其表達強度依次減弱。提示誘導和增強端粒酶反轉錄酶的表達對維持表皮幹細胞在體外自我更新和增殖能力可能具有重要意義。
