
物種學史
 獨角仙
獨角仙雙叉犀金龜最早是Linnaeus在1771年命名的,但林奈使用的屬名是 Scarabaeus,誤認為這是一種蜣螂。1911年Arrow以雲頂兜( Allomyrina pfeifferi)為模式種建立了 Allomyrina屬,並把戴叉犀金龜和雙叉犀金龜也置於此屬。而Minck在1920年又以雙叉犀金龜為模式標本建立了 Trypoxylus屬,因此雙叉犀金龜的屬名一直存在爭議。近年來染色體分析結果表明兩種叉犀金龜和雲頂兜的親緣關係較遠,因此作為獨立屬 Trypoxylus較為合適 。
形態特徵
成蟲
 雙叉犀金龜成蟲
雙叉犀金龜成蟲體長30-50mm(不含額角),體寬20-30mm,身體呈長卵圓形,體表背面光滑或微茸毛,腹面茸毛較多。體色個體差異較大,從深紅到紅棕再到純黑均有,有時同一個體的頭胸和鞘翅顏色也會不同。頭部較小,複眼深紅褐色,觸角有10節,其中鰓片部由3節組成,上唇呈上翹的扁平鏟狀。鞘翅肩疣、端疣發達,縱肋僅約略可辨。臀板十分短闊,強烈隆拱。足粗壯,前足脛節外緣3齒,基齒遠離端部兩齒,足末端有鉤爪一對。
性二型現象顯著,雄性有發達的額角,長15-35mm,向前伸出,末端向上彎曲並分四叉(體型較小的個體可能僅有兩叉),前胸背板中央向前伸出一前胸背角,略向下彎曲,末端分兩叉。雌性額頭頂部僅有一小型隆起,無前胸背角,胸背板有Y字型淺凹。雄性背部較為光亮,雌性背部較為粗暗。雄性前足明顯長於雌蟲。
幼蟲
 幼蟲♂注意其腹部刻點
幼蟲♂注意其腹部刻點獨角仙幼蟲俗稱“蠐螬”“雞母蟲”,身體肥大,最大可長約10cm。頭部堅硬,密布刻點,正面為黑色,邊緣為紅褐色;咀嚼式口器,大顎強而有力;單眼退化僅留痕跡。身體其他部分較為柔軟,常呈C型彎曲,乳白色或米黃色,體表半透明,腹部末端可見消化道內深色內容物;兩側有9對氣門,氣門較大,開放性無瓣膜;體表有稀疏的剛毛;六肢短小,運動主要靠身體蠕動。
雌性幼蟲小於雄性,雌性幼蟲體重峰值一般在20-25g,而雄性可達30-40g,甚至更重。此外L3時期,雄性幼蟲腹部倒數第二節中央可見一V字形刻點,而雌性幼蟲無此特徵。
棲息環境
多棲息於常綠或落葉闊葉林中,因幼蟲依賴林下的腐葉土或朽木形成的腐殖質為食,因而對森林的原生性要求較高,尤其喜愛殼斗科植物,如青岡櫟。此外幼蟲也會在腐爛的草堆、木材廠木屑堆、蘑菇種植場的木屑培養基中生活。
生活習性
 雙叉犀金龜幼蟲
雙叉犀金龜幼蟲幼蟲在腐殖土或木屑堆內部生活,一般不會將自己暴露在外面。因氣門開放,所以必須生活在潮濕的環境中,不然很容易因身體快速失水死亡。幼蟲在食物堆中緩慢活動並取食,排出橢圓形略扁的糞粒。此種幼蟲較為溫順,可以群居,但密度過大食物不足也會發生互咬現象。觸角有嗅覺功能,如果附近有蛋白、脂肪等含量較高的食物,幼蟲會前往取食。如果環境溫濕度不足、透氣性不足或食物耗盡,幼蟲會鑽出地表。
幼蟲最適溫度為23-25℃,此溫度下進食最活躍,生長狀況最好;雖然可以忍耐30℃的高溫,但會對生長產生不利影響;20℃以下進食量減少,生長緩慢;10℃以下進入休眠狀態,基本不進食,體重會減少;可以短時間忍受0℃左右的低溫,但低於-2℃會造成死亡。
幼蟲天敵主要為鼴鼠、獾等會挖掘的捕食者,以及叩甲、土蜂等肉食性幼蟲,也容易被真菌寄生死亡。
 舔舐樹汁
舔舐樹汁自然狀態下成蟲在初夏開始出現,最多見於6-7月,到9月則大部分都會死亡。成蟲在20℃以上活動,白天多在落葉堆、土壤內等陰暗潮濕環境休息,黃昏開始活動活動,活動高峰期為日落後夜晚的前幾個小時,有趨光性。成蟲喜食樹木的汁液或熟透的水果,雖然為咀嚼式口器,但取食方式高度特化,先用鏟狀的上唇劃破樹皮或水果表皮,然後用毛刷狀的舌舔舐樹汁或水果,樹汁也會吸引來其他昆蟲一起取食。如果夜間沒有攝入足夠的食物,也會出現白天繼續趴在食物上進食的情況。
雄性好鬥,為了爭奪食物和交配權會用額角搏鬥。搏鬥的雙方會先上下晃動額角,收縮腹部發出發出“嘰——嘰——”聲示威,如果雙方均不退卻,它們會努力將額角插入對手的身體下方,將對手舉起掀翻。被舉起的一方即為失敗者,一般只要被舉起一次就會立即認輸逃走。
成蟲天敵主要為烏鴉、貉、野豬等森林中的訊息肉食或雜食動物。體表常附著有大量的蟎,但一般不危及生命。
分布範圍
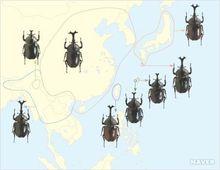 獨角仙分布範圍
獨角仙分布範圍雙叉犀金龜我國的主要分布區在秦嶺-淮河一線以南,北起陝西南部和河南南部,西起四川和陝西的整個南方地區,包括海南島和台灣島。記載上山東、河北、北京等華北地區也有分布,但近年來幾乎沒有發現記錄。但在吉林、遼寧 等東北地區卻有穩定分布,在朝鮮半島、琉球群島也有分布。雙叉犀金龜為什麼在我國呈現間斷分布有待進一步研究。
在日本分布於本州、四國、九州的一些島嶼,並因人為釋放擴散到了北海道。
在我國以南分布於越南、寮國、泰國的北部,緬甸、印度東部和孟加拉國。
繁殖方式
 獨角仙交配
獨角仙交配成熟後的雙叉犀金龜成蟲雄性有強烈的交配欲望,只要遇到雌性成蟲就會試圖交配。但是雌蟲對交配較為抗拒,特別是已經交配過的雌蟲,會拚命逃離雄蟲。雄蟲會用後足和中足攀附在樹皮上,前足抱住雌蟲進行交配。如果雌蟲正在進食,往往不會注意身後的雄蟲,這種情況下交配會比較順利。交配過程可以持續幾分鐘到幾十分鐘。交配(尤其是頻繁交配)會使雄蟲壽命縮短,因此在野外獨角仙雄性成蟲壽命僅有1個月左右,而雌性成蟲可達3個月,而人工飼養的獨角仙雄蟲如果不交配壽命也能達到3個月。
一般情況下,一次成功的交配就足夠雌蟲所有的卵受精。交配後幾天雌蟲就會開始產卵。雌蟲會尋找腐殖質堆等適合幼蟲生長的環境,鑽入食材堆中,把土壓緊,製造出一個比卵略大的卵室,每枚卵都有單獨的卵室。雌蟲在腐殖質中產卵一段時間後會出來覓食補充營養和體力,然後繼續尋找合適的場所產卵。一般情況下一隻雌蟲可以在2-3個多月的成蟲壽命里產下40-80枚卵。
 雙叉犀金龜蛹
雙叉犀金龜蛹卵乳白色或淡黃色,剛產下的卵性狀似大米粒,隨著發育會逐漸膨脹為綠豆粒大小的球形,溫濕度適宜的情況下孵化過程約為10天。剛孵化的幼蟲為一齡幼蟲,經過約1個月後蛻皮成為二齡幼蟲,再過1個月左右再次蛻皮成為三齡幼蟲。三齡幼蟲期最長,為6-8個月,隨生長環境溫度的變化會有延長和縮短。幼蟲的體重在三齡中期達到峰值,此時大概開始進入冬季,如果是人工條件下溫度恆定體重則基本保持不變;如果是自然條件下氣溫較低,幼蟲體重會明顯降低,到第二年氣溫變暖體重回升,但無法達到峰值水平。三齡末期的幼蟲停止進食,在土中用身體擠壓出一個橢球形的室,並用糞便和分泌物加固室壁,形成蛹室。雙叉犀金龜的蛹室豎直或微微傾斜,而大部分犀金龜的蛹室則是接近水平。此後幼蟲在蛹室內進入前蛹階段,身體變短,顏色變黃,表皮變皺,不久後蛻皮,即蛹化,蛹深黃色,沒有運動能力,只有腹部可以扭動。
雙叉犀金龜蛹經過20多天后羽化,剛羽化的成蟲鞘翅為白色而柔軟,經過幾小時後變為深色且硬化。剛羽化的成蟲不進食也不活動,會在蛹室中繼續蟄伏十幾天,然後從土中鑽出,開始覓食和交配。
所屬亞種
 T. d. takarai
T. d. takarai雙叉犀金龜主要有6-8個亞種:
T. d. dichotomus Linnaeus, 1771 分布於中國東部和南部
T. d. septentrionalis Kono, 1931 分布於日本本州,九州,四國
T. d. tunobosonis Kono, 1931 分布於中國台灣島
T. d. politus Prell, 1934 分布於泰國、緬甸、寮國和越南,印度東部
T. d. takarai Kusui, 1976 分布於日本沖繩本島
T. d. inchachina Kusui, 1976 分布於久米島,或與沖繩本島同一亞種
T. d. tuchiyai Nagai, 2006 分布於日本口永良部島
另外有 T. kanamorii Nagai, 2006 分布於緬甸北部一小塊區域,可能是獨立物種也可能是雙叉犀金龜的一個亞種。
此外近年來還有學者發布了 T. d. shennongji Takeuchi, 2014; T. d. xizangensis Li & Zhang, 2015 等新亞種 ,但其有效性還沒有被廣泛認可。
分布於中國東北和朝鮮半島的雙叉犀金龜亞種歸屬有爭議。
主要變種
將雙叉犀金龜作為寵物飼養由來已久,尤其是在日本。目前已經人工培育出了通體鮮紅色,紅色複眼或白色複眼的變種。
種群現狀
 被防鳥網纏死的雙叉犀金龜
被防鳥網纏死的雙叉犀金龜雙叉犀金龜目前不是珍稀物種,但是其生存也面臨一些威脅。
因為雙叉犀金龜可作為寵物飼養,且製成標本有較高的收藏觀賞價值,所以很多人在捕捉野生雙叉犀金龜。而且雙叉犀金龜成蟲有趨光性,很容易被吸引到危險的人類居住區。在果園人們布下的防鳥網很容易就會將獨角仙纏住造成大量死亡。
但是昆蟲有繁殖速度快且產卵量多的特點,所以人為捕捉並不是威脅雙叉犀金龜生存的主要原因。雙叉犀金龜幼蟲生長離不開森林下的腐殖質,所以對年代悠久的天然林非常依賴,在種植年代短的人工林中無法生存,所以對天然林的砍伐和破壞才是雙叉犀金龜面臨的最大威脅。
保護級別
雙叉犀金龜被列入《國家保護的有益的或者有重要經濟、科學研究價值的陸生野生動物名錄》(簡稱:三有名錄) ,受到《中華人民共和國野生動物保護法》的保護,不能隨意捕捉。
