
形態特徵
羽鰓類全部營固著生活,構成群體。這兩群的共同點是:原腔動物,身體分前、中、後三部分。在腸鰓類中稱為吻、領和軀幹,在羽鰓類中稱為頭盤、頸部和軀幹,都有1個或1對真體腔。腸鰓類消化道是直的,肛門開口在軀幹部的末端,在咽頭的側面排列著許多鰓裂。羽鰓類與營固著和群體生活相適應,消化道彎曲成V形,肛門位於頭盤的後方。在這兩群動物消化道前端的背面都有1個小盲囊(stomochord)有人認為它與脊索相同,而應歸入原索動物類,但無論是從組織學還是從發生學上來看,它都與脊索無關。有人認為半索動物與須腕動物和棘皮動物親緣關係較近,考慮與原索動物近似也有稱為前索動物(Prechordata)的。在庫根塔爾(W.Kükenthal)和克魯姆巴(T.Krumba-ch)的分類系統中,腸鰓類和羽鰓類被合稱為鰓孔類(Branchiotrema)。
 半索動物
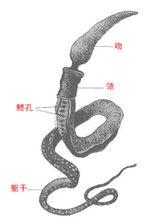
半索動物半索動物門(Hemichordata)動物,一類小型海產無脊椎動物,包括腸鰓類、羽鰓類(pterobranch)以及一屬只知道其幼蟲的浮球蟲屬(Planctosphaera)。前兩類的身體分成三部分;吻(管狀器官,在腸鰓類的吻通常發達)、領(羽鰓類的領有觸手)、軀幹(形長,通常又分數區)。半索動物用吻(腸鰓類)或觸手(羽鰓類)濾食水中漂浮的浮游生物,觸手上有纖毛和許多分泌細胞。浮球蟲的幼蟲生活于海洋表面,靠纖毛的運動引起水流,而攝食微小生物。大多數半索動物的生活史中有一幼蟲期,幼蟲稱柱頭幼蟲(參閱柱頭蟲〔acornworm〕條)。柱頭幼蟲和海星的羽腕幼蟲相似。單體或群體。無中樞神經系統,但領部有集中的神經組織。循環系統通常有可收縮的心囊、血管和血竇。咽部有多對鰓裂或無鰓裂。領部可有二或更多個具觸手的羽腕。雌雄異體,卵巢或精巢囊狀,沿軀幹前部排列成行。每囊有一孔開口到體表。半索動物與棘皮動物有密切的親緣關係,與棘皮動物及其他原索動物有共同的營固著生活或半固著生活的始祖。
綱目分類
半索門分三綱。腸鰓綱(Enteropneusta)常見於潮間帶和400公尺(約1,300呎)或更深的近海中。單體、蠕蟲狀、兩側對稱,鰓裂多數。巴西沿岸的大柱頭蟲(Balanoglossus gigas)體長達2公尺(約7呎)或更長。羽鰓綱(Pterobranchia)鰓裂有或無,領部具2個或更多的羽腕,聚居或群居於公共包被內。多種種類生活在600公尺(1,950呎)以下的近海,體微小、成管形。扁球綱(Planctosphaeroidea)僅發現一個種Planctosphaera pelagica的少數漂浮的幼體,透明,圓球形,兩側對稱,體表有曲折分支的纖毛帶。腸鰓綱體外受精,有些種類的受精卵發育為自由游泳的柱頭幼蟲,藉纖毛的運動作螺鏇式的運動。幼蟲可漂流達數周之久。以浮游生物為食,經變態發育為成體。僅知Balanoglossus屬一種柱頭蟲營無性生殖,由身體後端分離的小片段再生為成體。羽鰓綱的頭盤蟲屬(Cephalodiscus)及桿壁蟲屬(Rhabdopleura)的幼蟲以卵黃為養料,在其親體的公共包被內發育,後離親體入海水中,經短暫的自由游泳階段很快發育為成體。
腸鰓類通常生活在U形洞道內,大柱頭蟲的洞道長75公分(30吋)或更長。成體以吻部的張縮及領部的伸張和擴大鑽洞。體表具黏液,有潤滑、清潔和保護作用。鑽孔時將水及泥沙攝入口內,水中含有氧和食物微粒,後者在水流經鰓裂時被濾獲。
存在爭議
半索動物屬於在分類上有爭議的一個類群.
種類數量
全世界現生種不到100種,均為海生,營底棲生活。
代表動物
代表動物——柱頭蟲
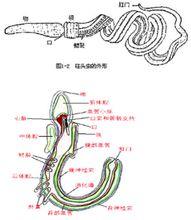
(一)有兩條縱走的神經索
有一條腹神經索和一條背神經索.背神經索在"領"中的部分有空腔,相當於背神經管的雛形).
(二)消化管前端咽壁的背側有許多對鰓裂.
(三)口上方向前伸出一條短盲管——口索.
(四)柱頭蟲其他特徵:
⒈棲息於太平洋沿岸淺海泥沙中,生活方式與蚯蚓相似.最長的個體可以超過兩尺.
⒉雌雄異體.變態發育,幼體與棘皮動物相似.
是介於脊索動物與棘皮動物之間的過渡類型.
物種起源
口索的起源與分類地位的確定:
(一)有學者認為口索可能是最初出現的脊索,因此認為半索動物屬於脊索動物門的一個亞門.
(二)有學者認為口索不是脊索的同源器官,口索很可能是內分泌系統的一個器官.而且柱頭蟲具有許多無脊椎動物的特徵,如具有腹神經索,開放式循環系統,肛在身體末端等特點.因此認為半索動物的分類地位應該是位於無脊椎動物與脊索動物之間的一個獨立的"門".
 半索動物
半索動物