無性生殖—多見於低等的無脊椎動物
在動物界,無性生殖幾乎僅見於一些低等的無脊椎動物(特別是在水或濕潤環境之中),雖然脊索動物中的尾索類也能進行無性生殖。無性生殖被分為以下類型:
裂殖(fission)
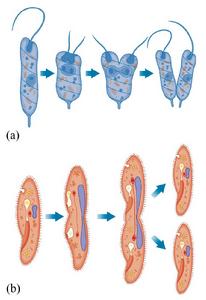 圖1原生動物的裂殖
圖1原生動物的裂殖動物身體直接進行分裂,分裂後每一部分都成為一個完整新個體。常見單細胞動物(原生動物),如草履蟲 的親體大致均等地分裂為兩個新個體(圖1);腔腸動物中的海葵也進行橫裂式或縱裂式的無性生殖;
出芽生殖(budding)
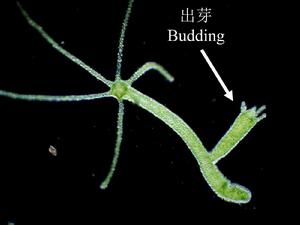 圖2 水螅的出芽生殖
圖2 水螅的出芽生殖從母體上長出芽,由芽發育成新個體的生殖方式(圖2),分為外出芽和內出芽。外出芽指開始時從親體上生出小突起,後經分化和長大而成為群體中的一員,有的脫離母體,成為獨立新個體,見於原生、多孔、腔腸、蠕蟲和尾索等類動物。殼吸管蟲的內出芽生殖是蟲體局部向內凹入以形成胚體,這時親體中大小核都進行分裂,所生子核進入胚體內,由此胚體長成的具纖毛幼蟲脫離母體後,在水中遊動,經變態後成為營附著生活的成體;淡水海綿以芽球萌發進行內出芽生殖。
斷裂生殖(fragmentation)
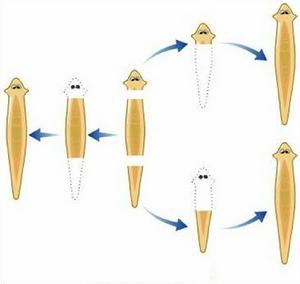 圖3 真渦蟲屬的斷裂生殖
圖3 真渦蟲屬的斷裂生殖沿動物身體主軸橫斷為兩部或多部,然後由各部分發育成新個體的方式(圖3),見於扁形動物中的單腸類和環節動物中的多毛類及寡毛類。
孤雌生殖(parthenogenesis reproduction)
也稱單性生殖,即卵不經過受精也能發育成正常的新個體的生殖方式。孤雌生殖常見於一些較原始的動物類群,如陸生無脊椎動物—蚜蟲、水生無脊椎動物—輪蟲、枝角類等。在這些類群中,無性生殖是主要的生殖方式,只有當惡劣的環境條件來臨時,才由孤雌生殖的雌蟲產生出雄蟲,進行兩性交配,產下休眠卵以度過嚴酷的條件。
無脊椎動物的出芽或斷裂生殖在機制上類似於高等植物的營養繁殖,但這類生殖在動物中其實十分罕見,相比之下,高等植物中營養繁殖要常見的多。為什麼會存在這種差異?這或許與動植物不同的結構發育以及防禦策略有關:“多細胞植物依靠由細胞壁構成的支架來支撐,這種骨架遍布全身。植物能在生命的全過程中,不斷生長和發育出新的部分……因為植株中每一個新的生長部分,都帶有用細胞壁來支撐的內部骨架,與此相反,動物用骨骼或堅韌的外殼來支撐,它們都由體內特定部位的特化細胞產生。發育過程大都集中在動物生命的早期階段,此時動物的所有組成部分均已成形……動物能通過奔跑、躲藏或格鬥來保護自己,植物則幾乎沒有逃避傷害的能力,但是,他能夠承受其軀體的大量損毀,因為植物有持續生長的能力”。因此,超強的再生與營養繁殖能力或許是植物在漫長的進化歷程中形成的一種生存策略,這既可以抵禦損傷的危害,又能快速擴張種群。
有性生殖—從接合生殖到融合生殖
有性生殖方式普遍見於各種動物類群,無論是低等的還是高等的。而一些高等動物只能進行有性生殖了。
接合生殖(conjugation)
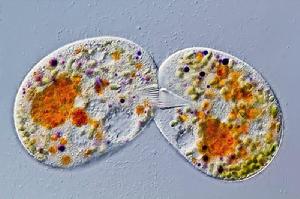 圖4 纖毛蟲的結合生殖
圖4 纖毛蟲的結合生殖指單細胞生物有性生殖由個體直接進行的生殖方式。結合生殖是一種較低級的有性生殖方式,常見於纖毛蟲(圖4),一般是兩個蟲體在胞口處互相連線,結合處胞膜消失,經過各自體內的核分裂並互相交換後,兩者又分 離,繼續進行二分裂形成新個體。如果接合子形態相同,稱為同配結合;如果接合子形態不同,稱為異配接合。
融合生殖(syngamy)
多細胞生物及單細胞生物的群體則由特化的單倍體細胞,即配子,進行融合生殖。融合生殖被區分為以下三種類型(圖5 a-c):
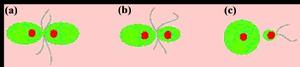 圖5 不同類型的融合生殖
圖5 不同類型的融合生殖1) 同配生殖(isogamy):配子的形態和機能完全相同,沒有性的區分,這可能是最原始的有性生殖; 2)異配生殖(heterogamy):一種是生理的異配生殖,參加結合的配子形態上並無區別,但交配型不同,只有不同交配型的配子才能結合;另一種是形態的異配生殖,參加結合的配子形狀相同,但大小和性表現不同,一般雌配子個體較大,不能運動,儲存了大量營養物質,而雄配子個體較小,有鞭毛,能通過遊動主動尋找卵子進行授精。譬如,人的卵要比精子大8500倍! 3)卵配生殖(oogamy):相結合的雌雄配子高度特化,其大小、形態和性表現都明顯不同,成為卵和精子。卵和精子經過受精,融合為受精卵。卵配生殖是分化顯著的異配生殖。
在有性生殖的早期,兩性配子在大小上差別不大。從進化的角度來看,融合生殖是朝著同配生殖→異配生殖→卵配生殖方向進化的,且雌配子越來越大。融合生殖是動物界最普遍的有性生殖方式,也廣泛存在於植物和真菌等類群。
有性生殖是所有高等脊椎動物的唯一生殖方式,雖然有報導罕見一些脊椎動物(如鳥類、蜥蜴、鯊魚等)在沒有受精的情況下也生下幼體,但這決不是普遍現象。因此,可以這樣說,在脊椎動物中,有性生殖被固化在生活史中,幾乎成為了生殖的唯一方式。這與植物顯然不同,如在最高等的植物—顯花植物中,無性的營養繁殖還很常見。
古爾德在分析性別的雙倍代價時推測“假設人類有這樣一個基因,它的正常型讓人類得以進行有性生殖,而它的突變性則導致女性可以進行單性生殖—生成可以在未受精狀態下自行發育的卵子(有些動物的確可以做到這一點,已經觀察到的蚜蟲、魚類以及蜥蜴)”,但是這種推測沒有任何實際意義,因為進化已經不再使哺乳動物存在無性生殖的生理基礎了,基於這樣的完全不可能存在的假設來談論性的代價已顯得十分荒唐。
雌雄配子的融合說起來簡單,但是動植物完成這一“性”行為的過程卻是千奇百怪,有些真是精彩迷人。讓我們來欣賞賈德森在 “動物性趣”中的一段精彩描述:“人類和許多動物會說性就是性交;青蛙和絕大多數的魚類會說性行為就是雙方在戰慄中排出卵子和精子;蠍子、千足蟲和蠑螈會告訴你,性行為就是一包包的精子排在地上,等待雌性坐上去後,精子包就會破裂,然後精子就會進入她的生殖道;海膽會說性行為就是在海水中排出卵子和精子,希望他們能在茫茫的海浪中找到對方;對於開花植物來說,性行為就是拜託風兒或者昆蟲將花粉捎給一朵等待中的雌花”。
單細胞生命體的繁殖通常就是一分為二,每一半都是完整的個體,但是在一些複雜的多細胞動植物(特別是不少的高等植物)中,卻依然留存了營養器官或組織的強有力的再生能力,“大自然使幾乎是獨立的性細胞具備了再生整個新個體的力量。但是,這種力量的某種東西卻依然分散於其餘的器官組織中,正如再生現象所證明的那樣,而可以想見:在某些特定情況下,這種機能可能整體地被保存在一種潛在狀態里,而一有機會就會顯示出來”。一方面,一個生殖細胞可以儲存個體發育的所有信息,一個營養細胞為什麼就不能?另一方面,這種再生應該具有快速的繁殖優勢,但為何未能廣泛擴散開來?推測可能存在其它的限制或選擇壓力,譬如,營養繁殖在水生高等植物中十分普遍,但是在乾燥的陸地上生存的高等動物幾乎沒有這種再生能力。
此外,在多細胞動物中偶爾也有單倍體個體存在,在這種情況下,減數分裂後的配子並不受精,而是通過繼續的細胞分裂直接發育成新個體,如雄蜂就是從蜂后的單倍體的卵產生的,它所有的體細胞都是單倍體。
性別二態性—在高等動物中可以極為誇張
“性”對動物形態的塑造程度似乎與進化程度密切相關。在原始的有性生殖中,雌體和雄體在形態結構上的差別往往難以區分,而隨著動物的進化,很多物種的雌體和雄體在形態結構上存在明顯的差異,即呈現出明顯的性別二態性(sexual dimorphism)。在一些高等動物中,性別二態性十分明顯,有些表現在體重上,有些則表現在形態、結構或顏色上。
 圖6 已經滅絕了的大角鹿
圖6 已經滅絕了的大角鹿為了在獲得與雌性交配權的戰鬥中取勝,在雄性個體之間殊死搏鬥的選擇壓力下,象海豹的雄性個體可比雌性個體重4-5倍,但是在交配時過重的雄性個體可壓得雌性個體窒息而死,因此自然選擇也會阻止雄性個體無限的 增大。大角鹿生活在距今300萬—1萬2千萬年前,亞歐大陸廣泛分布,以愛爾蘭地區的一種最著名,現已滅絕。大角鹿擁有極為誇張的鹿角(圖6),這種鹿角每年都會脫落,然後再重新長出,因此,雄鹿必須找到足夠的食物來供應每年更換角所需要消耗的能量,這樣,強大的角就是雄性活力、健康和武力的象徵。
還有許多動物通過展示華麗的魅力來爭奪與雌性的交配權,這種演化的一個極致就是百鳥之王的孔雀,與雌孔雀短短的尾巴相比,雄孔雀一到繁殖季節就撐開它那長長的尾巴展示一個五彩繽紛的巨大尾屏,跳起各種優美的舞蹈,以贏得雌孔雀的芳心,但是,雄孔雀的這個色澤艷麗的長尾巴其實也是個累贅,每年還得長出一些新羽毛替代前一年脫落的舊羽,這尾巴也不能成為攻擊或防禦的武器,若碰上獵食者,還會影響自己逃跑的速度,當然,如果這個炫耀的尾巴過大到顯著傷害其生存,自然的選擇就會適可而止。
性是如此的重要、有趣且魅力無窮—“如果不是為了性,大自然中絕大多數艷麗、漂亮的東西將不復存在:植物不會綻放花朵,鳥兒不會啾唧歌唱,鹿兒不再萌發鹿角,心兒也不會怦怦亂跳”。毫無疑問,性的自然選擇是生命進化的重要推動力之一。
雌雄同體—在動物中稀少,主要見於水生動物
雌雄同體(hermaphroditism)指兩性的機能或性狀在同一個個體中出現的現象。有兩種情況,一種是雌雄機能或性狀在一個個體上同時出現(固定型雌雄同體),另一種是一個個體可在雌雄性之間轉化,即可進行性逆轉(轉化型雌雄同體)。絕大多數被子植物都是雌雄同株(這其中絕大多數還是雌雄同花),與此不同的是,在動物中雌雄同體的現象還是比較稀少的。
 圖7 蚯蚓的交配
圖7 蚯蚓的交配軟體動物門腹足綱的後鰓亞綱動物都是固定性雌雄同體,在交配時,一對後鰓動物會互相貼近對方,然後交換精子。藤壺(節肢動物門顎足綱無柄目藤壺科)也是雌雄同體,但需異體受精。由於它們固著不能行動,在生 殖期間,必須靠著能伸縮的細管,將精子送入別的藤壺中使卵受精。蚯蚓是一種寡毛類動物,它們雌雄同體,但需要異體受精,交配時,兩條蚯蚓互抱,各排出精子輸入對方的受精囊內(圖7)。
許多海洋軟體動物(如牡蠣)在生命開始時是相對較小的雄性個體,隨著生長逐漸轉變為雌性個體,有時在生長過程中還會出現一段雌雄同體的時期(如帽貝)。在一些魚類的生活史中,性逆轉是一件經常發生的事,有的從雄性變為雌性(如小丑魚),而更常見的是雌性變為雄性,所有在珊瑚群中居住著的耀眼的魚類都採取這種策略(如隆頭魚),這種從雌魚向雄魚的轉變還見於“清潔魚”、海鱸、黃鱔等。
能夠進行性逆轉的多為水生動物,而在爬行動物、鳥類和哺乳動物中沒有發現過性逆轉的真實例子,這或許是因為為了適應陸地生存和繁殖的需要,陸生動物進化出了一系列更為複雜的受精和卵保護技術,以防止卵的水分蒸發,與這個特殊的發展相稱的是,雌性個體在解剖學上更加雌性化,具有特殊的導管和腺體,使卵外包被防水的殼,並且最後進化出可供幼體在其體內發育的子宮,雌性的這種日益完善的結構更深刻地影響著她們與雄性之間的關係,而雄性則向身體結構更適合角斗和求愛方向發展,這樣兩性沿著不可逆的方向不斷進化與發展。
如果性別的雙倍代價對動物成立的話,那在動物中這種雌雄同體現象就應該相當普遍,可實際情況卻正好相反。
