軟骨魚綱(化石)
正文
魚類的一大類群,一般列為綱,也有人列為亞綱,置於廣義魚綱(Pisces)中,具有軟骨質內骨骼,體被盾鱗,雄性有交接器,無鰾。內骨骼可鈣化成獨特的稜柱顆粒表層。外骨骼由小鱗組成,有時擴大(或癒合)成頭棘或鰭棘,通常很少形成甲片。大多數種類的鰭膜有角質鰭條支持。舌弓鰓裂常退化成噴水孔或消失。舌頜骨左頜懸掛方面起著重要作用。有直接開口於體外的鰓裂 5至7對,在全頭類有4對,外被有一膜質鰓蓋。嗅囊相當大,嗅覺靈敏。吻突由內吻軟骨支持。口腹位,鼻孔也或多或少居腹位,並可與口匯合,鼻孔被垂瓣完全地分為入水孔和出水孔。牙齒不與頜骨癒合,並新舊依次替換。鰓弓一般呈“>”形,而不象硬骨魚類的“∑”形,但均有例外。側線管穿過兩側鱗片之間,並在頭部與壺腹感覺器群相連。迷路通過內淋巴管開向外方,板鰓類者內有耳砂(平衡錐)。古生代軟骨魚類的鱗片在形狀和構造上變化很大,與晚期種類的盾鱗有所不同,如晚石炭世的全中鯊(Ho-lmesella) 的鱗是典型的複合生長結構。研究這種結構的生成過程的稱鱗粒理論,即鱗粒是鱗發生中的最小單位,常常是若干鱗粒癒合成大的鱗片,鱗粒癒合方式不同而有圓鱗粒鱗和同時鱗粒鱗之分,然而也可以出現很簡單的鱗粒單位存在。較晚期的軟骨類的盾鱗是不生長的同時鱗粒鱗。即在盾鱗中各個鱗粒是同時出現的,並且是在齒質冠形成之前的乳突階段癒合的,結果則使鱗的發育總時間縮短了。隨著鱗粒癒合的程度情況,可以有數個不完全分開的髓腔,或者只有一個大髓腔。
由於軟骨魚類的骨骼為軟骨,在地層中除保存有牙齒和棘刺外,其他部分的化石極為稀少,而牙齒又多呈分散狀態保存,難於了解其排列組合形式。許多古生代的屬種僅是以個別牙齒或殘片為代表,從形態上進行分類(圖1),故對一些主要類群的發展歷史及彼此間的親緣關係仍不清楚。
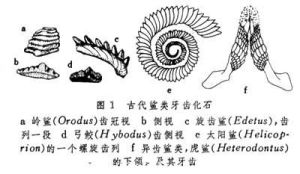 軟骨魚綱(化石)
軟骨魚綱(化石)軟骨魚類的進化可分為兩個方向:即板鰓類方向和全頭類方向,且兩者早就各自分別地發展。一般將板鰓類的歷史分為3個階段:①原始的裂口鯊階段主要生存於泥盆紀,延續到晚古生代;②弓鮫階段約始自早石炭世,到三疊紀仍相當繁盛;③近世階段自侏羅紀始,輻射發展出後來的鯊類及其親族。然而 3階段並不是銜接的直接關係。板鰓類的分類概述如下:
板鰓亞綱
裂口鯊目,中泥盆世-早二疊世(圖2):
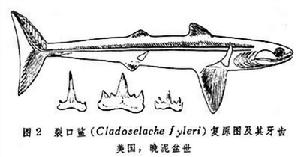 軟骨魚綱(化石)
軟骨魚綱(化石)異棘鯊目,晚泥盆世-中二疊世
太陽鯊目,晚石炭世-早三疊世
鯊目(=Euselachii)
櫛棘鯊亞目,晚泥盆世-晚石炭世
弓鮫亞目,早石炭世-晚白堊世
虎鯊亞目,早侏羅世-現代
六鰓鯊亞目,早侏羅世-現代
鼠鯊亞目,晚侏羅世-現代
貓鯊亞目,晚侏羅世-現代
角鯊亞目,晚侏羅世-現代
扁鯊亞目,晚侏羅世-現代
鋸鯊亞目,晚白堊世-現代
鰩目
犁頭鰩亞目,早侏羅世-現代
鋸鰩亞目,早白堊世-現代
亞目,早白堊世-現代
鰩亞目,晚白堊世-現代
鱝亞目,晚白堊世-現代
電鰩亞目,古新世-現代
軟骨魚類的第二條進化路線以全頭類為代表。這可從現代的銀鮫類經由中生代的多棘鮫類追溯到頰甲鮫類。它們幾乎全是底棲的,具有替換緩慢的齒板,基本以帶殼食物為食,用齒板研磨。全頭類於石炭紀達到極盛期,侵占了原來被盾皮魚類占據的環境,並取而代之。全頭類中有兩類齒型:即“鯊”型和“齒板”型。在“鯊”型中有像在板鰓類中那樣分開的連續牙齒;而在“齒板”型中,牙齒癒合成替換緩慢的研磨齒板。一般分類如下:
全頭亞綱
軟鱔鮫目,早石炭世
旋齒鯊亞目,早石炭世-早三疊世
砂齒鯊亞目,早石炭世
寇氏鯊目,晚泥盆世-晚石炭世
瓣齒鯊目,早石炭世-晚二疊世
銀鮫目
單疣齒鮫亞目,晚泥盆世-二疊紀
殼齒鮫亞目,石炭紀
頰甲鮫亞目,石炭紀-晚二疊世
多棘鮫亞目,侏羅紀
似鮫亞目,早侏羅世
銀鮫亞目,早侏羅世-現代
過去曾有人將瓣齒類、殼齒類、砂齒類和寇氏鯊類歸為一類,並由於它們的牙齒均替換緩慢,而名為緩齒魚類,近來已被廢棄。它們分別獨立為目,可能更便於探討它們的分類位置。
一般對板鰓類和全頭類做為一個自然類群是無爭議的。也有人主張用板鰓形類(Elasmobranchiomorpha)這一名詞來總括起板鰓類、全頭類及繁盛於泥盆紀的盾皮魚類;並認為前兩類是在由板鰓形類基幹上分化出盾皮魚類和軟骨魚類兩大支後,才由後者再行分支的。
